Строение и функции ядра
Ядро — важнейший органоид клетки, характерный для эукариотов и являющийся признаком высокой организации организма. Ядро является центральным органоидом. Оно состоит из ядерной оболочки, кариоплазмы (ядерной плазмы), одного или нескольких ядрышек (у некоторых организмов ядрышки в ядре отсутствуют); в состоянии деления возникают особые органоиды ядра — хромосомы.
1. Ядерная оболочка.
Строение ядерной оболочки аналогично таковому для клеточной мембраны. Она содержит поры, осуществляющие контакт содержимого ядра и цитоплазмы.
Функции ядерной оболочки :
1) отделяет ядро от цитоплазмы;
2) осуществляет взаимосвязь ядра и остальных органоидов клетки.
2. Кариоплазма (ядерная плазма).
Кариоплазма представляет собой жидкий коллоидно-истинный раствор, содержащий белки, углеводы, соли, другие органические и неорганические вещества. В кариоплазме содержатся все нуклеиновые кислоты: практически весь запас ДНК, информационные, транспортные и рибосомальные РНК. Строение кариоплазмы зависит от функционального состояния клетки. Функциональных состояний клетки эукариотов два: стационарное и состояние деления.
В стационарном состоянии (это или время между делениями, т. е. интерфаза, или время обычной жизнедеятельности специализированной клетки в организме) нуклеиновые кислоты равномерно распределены в кариоплазме, ДНК — деспирализованы и структурно не выделены. В ядре нет других органоидов, кроме ядрышек (если таковые характерны для данной клетки), ядерной оболочки и кариоплазмы.
В состоянии деления ядерные кислоты образуют особые органоиды — хромосомы, ядерное вещество становится хроматиновым (способным к окрашиванию). В процессе деления ядерная оболочка растворяется, ядрышки исчезают, а кариоплазма смешивается с цитоплазмой.
Хромосомы представляют собой особые образования определенной формы. По форме различают палочкообразные, разноплечные и равноплечные хромосомы, а также хромосомы с вторичными перетяжками. Тело хромосомы состоит из центромеры и двух плеч.
У палочкообразных хромосом одно плечо очень большое, а второе — маленькое, у разноплечных — оба плеча соизмеримы друг с другом, но видимо различаются по размерам, у равноплечных размеры плеч одинаковы.
Число хромосом для каждого вида строго одинаково и является систематическим признаком. Известно, что в многоклеточных организмах различают два типа клеток по количеству хромосом — соматические (клетки тела) и половые клетки, или гаметы. Число хромосом в соматических клетках (в норме, как правило) в два раза больше, чем в половых клетках. Поэтому число хромосом в соматических клетках называют диплоидным (двойным), а количество хромосом в гаметах — гаплоидным (одинарным). Например, в соматических клетках тела человека содержится 46 хромосом, т. е. 23 пары (это диплоидный набор); половые клетки человека (яйцеклетки и сперматозоиды) содержат 23 хромосомы (гаплоидный набор).
Парные хромосомы имеют одинаковую форму и выполняют одинаковые функции: они несут информацию об одинаковых типах признаков (например, половые хромосомы несут информацию о поле будущего организма).
Парные хромосомы, имеющие одинаковое строение и выполняющие одинаковые функции, называются аллельными (гомологичными).
Хромосомы, принадлежащие к разным парам гомологичных хромосом, называются неаллельными.
Диплоидный набор хромосом обозначается «2n», а гаплоидный — «n»; следовательно, в соматических клетках содержится 2n хромосом, а в гаметах — n хромосом.
Число хромосом в клетке не является показателем уровня организации организма (дрозофила, принадлежащая к насекомым — организмам высокого уровня организации, — содержит в соматических клетках четыре хромосомы).
Хромосомы состоят из генов.
Ген — участок молекулы ДНК, в котором закодирован определенный состав молекулы белка, за счет чего у организма проявляется тот или иной признак, или реализующийся у конкретного организма, или передающийся от родительского организма потомкам.
Итак, хромосомы — это органоиды, которые четко проявляются в клетках в момент деления последних. Они образованы нуклеопротеидами и выполняют в клетке следующие функции:
1) хромосомы содержат наследственную информацию о признаках, присущих данному организму;
2) через хромосомы осуществляется передача наследственной информации потомству.
3. Ядрышко.
Небольшое сферическое образование, содержащееся внутри кариоплазмы, называется ядрышком. В ядре может содержаться одно или несколько ядрышек, но ядрышко может и отсутствовать. В ядрышке более высокая концентрация матрикса, чем в кариоплазме. Оно содержит различные белки, в том числе и нуклеопротеиды, липопротеиды, фосфопротеиды.
Главной функцией ядрышек является синтез зародышей рибосом, которые сначала попадают в кариоплазму, а затем через поры в ядерной оболочке — в цитоплазму на эндоплазматическую сеть.
4. Общие функции ядра:
1) в ядре сосредоточена практически вся информация о наследственных признаках данного организма (информативная функция);
2) ядро через гены, содержащиеся в хромосомах, передает признаки организма от родителей к потомкам (функция наследования);
3) ядро является центром, объединяющим все органоиды клетки в единое целое (функция объединения);
4) ядро согласует и регулирует физиологические процессы и биохимические реакции в клетках (функция регуляции).
Лекция № .
Количество часов: 2
Клеточное ЯДРО
1. Общая характеристика интерфазного ядра. Функции ядра
2.
3.
4.
1. Общая характеристика интерфазного ядра
Ядро - это важнейшая составная часть клетки, которая имеется практически во всех клетках многоклеточных организмов. Большинство клеток имеет одно ядро, но бывают двуядерные и многоядерные клетки (например, поперечно-полосатые мышечные волокна). Двуядерность и многоядерность обусловлены функциональными особенностями или патологическим состоянием клеток. Форма и размеры ядра очень изменчивы и зависят от вида организма, типа, возраста и функционального состояния клетки. В среднем объем ядра составляет приблизительно 10% от общего объема клетки. Чаще всего ядро имеет округлую или овальную форму размером от 3 до 10 мкм в диаметре. Минимальный размер ядра составляет 1 мкм (у некоторых простейших), максимальный - 1 мм (яйцеклетки некоторых рыб и земноводных). В некоторых случаях наблюдается зависимость формы ядра от формы клетки. Ядро обычно занимает центральное положение, но в дифференцированных клетках может быть смещено к периферийному участку клетки. В ядре сосредоточена практически вся ДНК эукариотической клетки.
Основными функциями ядра являются:
1) Хранение и передача генетической информации;
2) Регуляция синтеза белка, обмена веществ и энергии в клетке.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводится. Поэтому нарушение любой из этих функций приведет к гибели клетки. Все это указывает на ведущее значение ядерных структур в процессах синтеза нуклеиновых кислот и белков.
Одним из первых ученых продемонстрировавших роль ядра в жизнедеятельности клетки был немецкий биолог Хаммерлинг. В качестве экспериментального объекта Хаммерлинг использовал крупные одноклеточные морские водоросли Acetobularia mediterranea и А. c renulata. Эти близкородственные виды хорошо отличаются друг от друга по форме «шляпки». В основании стебелька находится ядро. В одних экспериментах шляпку отделяли от нижней части стебелька. В результате было установлено, что для нормального развития шляпки необходимо ядро. В других экспериментах стебелек с ядром одного вида водоросли соединялся со стебельком без ядра другого вида. У образовавшихся химер всегда развивалась шляпка, типичная для того вида, которому принадлежало ядро.
Общий план строения интерфазного ядра одинаков у всех клеток. Ядро состоит из ядерной оболочки, хроматина, ядрышек, ядерного белкового матрикса и кариоплазмы (нуклеоплазмы). Эти компоненты встречаются практически во всех неделящихся клетках эукариотических одно- и многоклеточных организмов.
2. Ядерная оболочка, строение и функциональное значение
Ядерная оболочка (кариолемма, кариотека) состоит из внешней и внутренней ядерных мембран толщиной по 7 нм. Между ними располагается перинуклеарное пространство шириной от 20 до 40 нм. Основными химическими компонентами ядерной оболочки являются липиды (13-35%) и белки (50-75%). В составе ядерных оболочек обнаруживаются также небольшие количества ДНК (0-8%) и РНК (3-9%). Ядерные оболочки характеризуются относительно низким содержанием холестерина и высоким - фосфолипидов. Ядерная оболочка непосредственно связана с эндоплазматической сетью и содержимым ядра. С обеих сторон к ней прилегают сетеподобные структуры. Сетеподобная структура, выстилающая внутреннюю ядерную мембрану, имеет вид тонкой оболочки и называется ядерной ламиной. Ядерная ламина поддерживает мембрану и контактирует с хромосомами и ядерными РНК. Сетеподобная структура, окружающая наружную ядерную мембрану, гораздо менее компактна. Внешняя ядерная мембрана усеяна рибосомами, участвующими в синтезе белка. В ядерной оболочке имеются многочисленные поры диаметром около 30-100 нм. Количество ядерных пор зависит от типа клетки, стадии клеточного цикла и конкретной гормональной ситуации. Так чем интенсивнее синтетические процессы в клетке, тем больше пор имеется в ядерной оболочке. Ядерные поры довольно лабильные структуры, т. е. в зависимости от внешнего воздействия способны изменять свой радиус и проводимость. Отверстие поры заполнено сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют ядерным поровым комплексом. Сложный комплекс пор имеет октагональную симметрию. По границе округлого отверстия в ядерной оболочке располагаются три ряда гранул, по 8 штук в каждом: один ряд лежит средство построения концептуальных моделей стороны ядра, другой - средство построения концептуальных моделей стороны цитоплазмы, третий расположен в центральной части пор. Размер гранул около 25 нм. От гранул отходят фибриллярные отростки. Такие фибриллы, отходящие от периферических гранул, могут сходиться в центре и создавать как бы перегородку, диафрагму, поперек поры. В центре отверстия часто можно видеть так называемую центральную гранулу.
Ядерно-цитоплазматический транспорт
Процесс транслокации субстрата через ядерную пору (для случая импорта) состоит из нескольких стадий. На первой стадии транспортирующийся комплекс заякоривается на обращенной в цитоплазму фибрилле. Затем фибрилла сгибается и перемещает комплекс ко входу в канал ядерной поры. Происходит собственно транслокация и освобождение комплекса в кариоплазму. Известен и обратный процесс - перенос веществ из ядра в цитоплазму. Это в первую очередь касается транспорта РНК синтезируемого исключительно в ядре. Также существует другой путь переноса веществ из ядра в цитоплазму. Он связан с образованием выростов ядерной оболочки, которые могут отделяться от ядра в виде вакуолей, а затем содержимое их изливается или выбрасывается в цитоплазму.
Таким образом, обмен веществ между ядром и цитоплазмой осуществляется двумя основными путями: через поры и путем отшнуровывания.
Функции ядерной оболочки:
1. Барьерная. Эта функция заключается в отделении содержимого ядра от цитоплазмы. В результате оказываются пространственно разобщенными процессы синтеза РНК/ДНК от синтеза белка.
2. Транспортная. Ядерная оболочка активно регулирует транспорт макромолекул между ядром и цитоплазмой.
3. Организующая. Одной из основных функций ядерной оболочки является ее участие в создании внутриядерного порядка.
3. Строение и функции хроматина и хромосом
Наследственный материал может находиться в ядре клетки в двух структурно-функциональных состояниях:
1. Хроматин. Это деконденсированное, метаболически активное состояние, предназначенное для обеспечения процессов транскрипции и редупликации в интерфазе.
2. Хромосомы. Это максимально конденсированное, компактное, метаболически неактивное состояние, предназначенное для распределения и транспортировки генетического материала в дочерние клетки.
Хроматин. В ядре клеток выявляются зоны плотного вещества, которые хорошо окрашиваются основными красителями. Эти структуры получили название "хроматин" (от греч. «хромо» – цвет, краска). Хроматин интерфазных ядер представляет собой хромосомы, находящиеся в деконденсированном состоянии. Степень деконденсации хромосом может быть различной. Зоны полной деконденсации называются эухроматином. При неполной деконденсации в интерфазном ядре видны участки конденсированного хроматина, называемого гетерохроматином. Степень деконденсации хроматина в интерфазе отражает функциональную нагрузку этой структуры. Чем "диффузнее" распределен хроматин в интерфазном ядре, тем интенсивнее в нем синтетические процессы. Уменьшение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина. Максимальная конденсация конденсированного хроматина достигается во время митотического деления клеток. В этот период хромосомы не выполняют никаких синтетических функций.
В химическом отношении хроматин состоит из ДНК (30-45%), гистонов (30-50%), негистонных белков (4-33%) и небольшого количества РНК. ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. Репликоны - участки ДНК, которые синтезируются как независимые единицы. Репликоны имеют начальную и терминальную точки синтеза ДНК. РНК представляет собой все известные клеточные типы РНК, находящиеся в процессе синтеза или созревания. Гистоны синтезируются на полисомах в цитоплазме, причем этот синтез начинается несколько раньше редупликации ДНК. Синтезированные гистоны мигрируют из цитоплазмы в ядро, где и связываются с участками ДНК.
В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, ассоциированной с гистонами. Хроматиновая нить представляет собой двойную спираль ДНК, окружающую гистоновый стержень. Она состоит из повторяющихся единиц – нуклеосом. Количество нуклеосом огромно.
Хромосомы (от. греч. хромо и сома) - это органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов.
Хромосомы представляют собой палочковидные структуры разной длины с довольно постоянной толщиной. У них имеется зона первичной перетяжки, которая делит хромосому на два плеча. Хромосомы с равными называют метацентрическими , с плечами неодинаковой длины - субметацентрическими. Хромосомы с очень коротким, почти незаметным вторым плечом называются акроцентрическими.
В области первичной перетяжки находится центромера, представляющая собой пластинчатую структуру в виде диска. К центромере прикрепляются пучки микротрубочек митотического веретена, идущие в направлении к центриолям. Эти пучки микротрубочек принимают участие в движении хромосом к полюсам клетки при митозе. Некоторые хромосомы имеют вторичную перетяжку. Последняя обычно расположена вблизи дистального конца хромосомы и отделяет маленький участок, спутник. Вторичные перетяжки называют ядрышковыми организаторами. Здесь локализована ДНК, ответственная за синтез р-РНК. Плечи хромосом оканчиваются теломерами, конечными участками. Теломерные концы хромосом не способны соединяться с другими хромосомами или их фрагментами. В отличие от них разорванные концы хромосом могут присоединяться к таким же разорванным концам других хромосом.
Размеры хромосом у разных организмов варьируют в широких пределах. Так, длина хромосом может колебаться от 0,2 до 50 мкм. Самые мелкие хромосомы обнаруживаются у некоторых простейших, грибов. Наиболее длинные - у некоторых прямокрылых насекомых, у амфибий и у лилейных. Длина хромосом человека находится в пределах 1,5-10 мкм.
Число хромосом у различных объектов также значительно колеблется, но характерно для каждого вида животных или растений. У некоторых радиолярий число хромосом достигает 1000-1600. Рекордсменом среди растений по числу хромосом (около 500) является папоротник ужовник, 308 хромосом у тутового дерева. Наименьшее количество хромосом (2 на диплоидный набор) наблюдается у малярийного плазмодия, лошадиной аскариды. У человека число хромосом составляет 46, у шимпанзе, таракана и перца – 48, плодовая мушка дрозофила – 8, домашняя муха – 12, сазана – 104, ели и сосны – 24, голубя - 80.
Кариотип (от греч. Карион - ядро, ядро ореха, операторы - образец, форма) - совокупность признаков хромосомного набора (число, размер, форма хромосом), характерные для того или иного вида.
Особи разного пола (особенно у животных) одного и того же вида могут различаться по числу хромосом (различие чаще всего на одну хромосому). Даже у близких видов хромосомные наборы отличаются друг от друга или по числу хромосом, или по величине хотя бы одной или нескольких хромосом. Следовательно, структура кариотипа может быть таксономическим признаком.
Во второй половине 20 века в практику хромосомного анализа стали внедряться методы дифференциального окрашивания хромосом. Считается, что способность отдельных участков хромосом к окрашиванию связана с их химическими различиями.
4. Ядрышко. Кариоплазма. Ядерный белковый матрикс
Ядрышко (нуклеола) - обязательный компонент клеточного ядра эукариотных организмов. Однако имеются некоторые исключения. Так ядрышки отсутствуют в высокоспециализированных клетках, в частности в некоторых клетках крови. Ядрышко представляет собой плотное тельце округлой формы величиной 1-5 мкм. В отличие от цитоплазматических органоидов ядрышко не имеет мембраны, которая окружала бы его содержимое. Размер ядрышка отражает степень его функциональной активности, которая широко варьирует в различных клетках. Ядрышко является производным хромосомы. В состав ядрышка входят белок, РНК и ДНК. Концентрация РНК в ядрышках всегда выше концентрации РНК в других компонентах клетки. Так концентрация РНК в ядрышке может быть в 2-8 раз выше, чем в ядре, и в 1-3 раза выше, чем в цитоплазме. Благодаря высокому содержанию РНК, ядрышки хорошо окрашиваются основными красителями. ДНК в ядрышке образует большие петли, которые носят название «ядрышковые организаторы». От них зависит образование и количество ядрышек в клетках. Ядрышко неоднородно по своему строению. В нем выявляются два основных компонента: гранулярный и фибриллярный. Диаметр гранул около 15-20 нм, толщина фибрилл – 6-8 нм. Фибриллярный компонент может быть сосредоточен в центральной части ядрышка, а гранулярный - по периферии. Часто гранулярный компонент образует нитчатые структуры - нуклеолонемы толщиной около 0, 2 мкм. Фибриллярный компонент ядрышек представляет собой рибонуклеопротеидные тяжи предшественников рибосом, а гранулы - созревающие субъединицы рибосом. Функция ядрышка заключается в образовании рибосомных РНК (рРНК) и рибосом, на которых происходит синтез полипептидных цепей в цитоплазме. Механизм образования рибосом следующий: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком. В зоне ядрышка происходит сборка субъединиц рибосом. В активно функционирующих ядрышках синтезируется 1500-3000 рибосом в минуту. Рибосомы из ядрышка через поры в ядерной оболочке поступают на мембраны эндоплазматической сети. Количество и образование ядрышек связано с активностью ядрышковых организаторов. Изменения числа ядрышек могут происходить за счет слияния ядрышек или при сдвигах в хромосомном балансе клетки. Обычно в ядрах содержится несколько ядрышек. В ядрах некоторых клеток (ооциты тритонов) содержится большое количество ядрышек. Это явление получило название амплификации. Оно заключается в организации систем управления качеством, что происходит сверхрепликация зоны ядрышкового организатора, многочисленные копии отходят от хромосом и становятся дополнительно работающими ядрышками. Такой процесс необходим для накопления огромного количества рибосом на яйцеклетку. Благодаря этому обеспечивается развитие эмбриона на ранних стадиях даже при отсутствии синтеза новых рибосом. Сверхчисленные ядрышки после созревания яйцевой клетки исчезают.
Судьба ядрышка при делении клеток. По мере затухания синтеза р-РНК в профазе происходит разрыхление ядрышка и выход готовых рибосом в кариоплазму, а затем и в цитоплазму. При конденсации хромосом фибриллярный компонент ядрышка и часть гранул тесно ассоциируют с их поверхностью, образуя основу матрикса митотических хромосом. Этот фибриллярно-гранулярный материал переносится хромосомами в дочерние клетки. В ранней телофазе по мере деконденсации хромосом происходит высвобождение компонентов матрикса. Его фибриллярная часть начинает собираться в мелкие многочисленные ассоциаты - предъядрышки, которые могут объединяться друг с другом. По мере возобновления синтеза РНК предъядрышки превращаются в нормально функционирующие ядрышки.
Кариоплазма (от греч. < карион > – орех, ядро ореха), или ядерный сок, в виде бесструктурной полужидкой массы окружает хроматин и ядрышки. Ядерный сок содержит белки и различные РНК.
Ядерный белковый матрикс (ядерный скелет) - каркасная внутриядерная система, которая служит для поддержания общей структуры интерфазного ядра объединения всех ядерных компонентов. Представляет собой нерастворимый материал, остающийся в ядре после биохимических экстракций. Он не имеет четкой морфологической структуры и состоит на 98% из белков.
Ядерная оболочка (нуклеолемма) представляет собой сложное образование, отделяющее содержимое ядра от цитоплазмы и других элементов живой клетки. Данная оболочка выполняет ряд важных функций, без которых является невозможным функционирование ядер, полноценное . Чтобы определить роль ядерных мембран в жизнедеятельности эукариотных клеток, необходимо узнать не только главные функции, но и особенности строения.
В статье подробно рассматриваются функции ядерной оболочки. Описывается строение, структурные компоненты нуклеолеммы, их взаимосвязь, механизмы транспортировки веществ, процесс деления при митозе.
Строение оболочки
Главное отличие эукариот заключается в наличии ядра и ряда других органелл, необходимых для его поддержания. Такие клетки входят в состав всех растений, грибов, животных, в то время как клетки-прокариоты представляют собой простейшие безъядерные организмы.
Нуклеолемма состоит из двух структурных элементов - внутренней и наружной мембран. В промежутке между ними существует свободное пространство, называемое перинуклеарным. Ширина перинуклеарного промежутка нуклеолеммы составляет от 20 до 60 нанометров (нм).
Внешняя мембрана нуклеолеммы контактирует с клеточной цитоплазмой. На ее наружной поверхности располагается существенное число рибосом, которые отвечают за из отдельных аминокислот. Внешняя мембрана не содержит рибосом.

Мембраны, образующие нуклеолемму, состоят из белковых соединений и двойного слоя фосфолипидных веществ. Механическая прочность оболочки обеспечивается сетью филаментов - нитевидных белковых структур. Наличие филаментной сети характерно для большинства эукариот. Они соприкасаются с внутренней мембраной.
Сети филаментов располагаются не только в области нукелолеммах. Такие структуры также располагаются в цитоплазме. Их функция заключается в сохранении целостности клетки, а также в формировании контактов между клетками. При этом, отмечается, что слои, образующие сеть, регулярно перестраиваются. Данный процесс наиболее активен в период роста клеточного ядра перед делением.
Сеть филаментов, которая поддерживает мембраны, называется ядерной ламиной. Она формируется из определенной последовательности белков-полимеров, которые называются ламинами. Она взаимодействует с хроматином - веществом, участвующим в формировании хромосом. Также ламина контактирует с молекулами рибонуклеиновой кислоты, ответственными за .
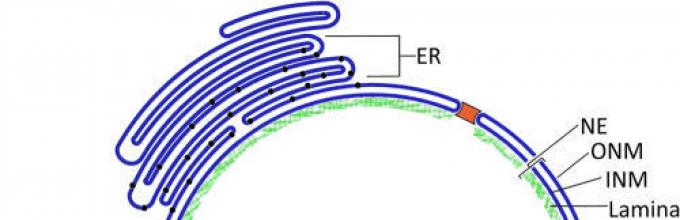
Внешняя мембрана ядра взаимодействует с мембраной, окружающей эндоплазматический ретикулум. В определенных участках оболочки происходит контакт перинуклеарного пространства и внутреннего пространства ретикулума.
Функции эндоплазматического ретикулума:
- Синтез и транспортировка белков
- Хранение продуктов синтеза
- Формирование новой оболочки при митозе
- Хранение , выполняющих функцию медиатора
- Продукция гормонов

Внутри оболочки располагаются ядерные поровые комплексы. Это каналы, посредством которых происходит перенос молекул между клеточным ядром, цитоплазмой и другими клеточными органеллами. На одном квадратном микроне поверхности нуклеолеммы располагает от 10 до 20 поровых комплекса. Исходя из этого, в оболочке 1 соматической клетки может находится всего от 2 до 4 тысяч ЯПК.
Помимо транспорта веществ, оболочка выполняет опорную и защитную функцию. Она отделяет ядро от содержимого цитоплазмы, в том числе продуктов деятельности других органелл. Защитная функция заключается в предохранении генетической информации ядра от негативного воздействия, например, .
Считается, что двойная мембрана ядерной оболочки сформировалась в ходе эволюции путем захвата одних клеток другими. Вследствие этого, некоторые поглощенные клетки сохранили собственную активность, но при этом их ядро было окружено двойной мембраной - собственной, и мембраной клетки-хозяина.
Таким образом, ядерная оболочка представляет собой сложную структуру, состоящую из двойной мембраны, содержащей ядерные поры.
Строение и свойства ЯПК
Ядерный поровый комплекс - это симметричный канал, местом локализации которого является сличение наружной и внутренней мембран. ЯПК состоят из набора веществ, включающих около 30 видов белков.
Ядерные поры имеют бочкообразную форму. Образуемый канал не ограничивается ядерными мембранами, а незначительно выступает за их пределы. В результате с двух сторон оболочки возникают кольцеобразные выступы. Размер этих выступов отличается, так как с одной стороны кольцеобразное образование имеет больший диаметр, чем с другой. Элементы ядерных пор, выступающих за пределы мембраны, называются терминальными структурами.
![]()
Цитоплазматическая терминальная структура (та что находится на внешней поверхности ядерной мембраны) состоит из восьми коротких фибрилл-нитей. Ядерная терминальная структура также состоит из 8 фибрилл, однако они образуют кольцо, выполняющего функции корзины. Во многих клетках от ядерной корзины исходят дополнительные фибриллы. Терминальные структуры являются местами, где происходит контакт молекул, транспортируемых через ядерные поры.
В месте расположения ЯПК происходит слияние наружной и внутренней ядерной мембраны. Такое слияние объясняется необходимостью обеспечить фиксацию ядерных пор в мембранах с помощью белков, соединяющих их также с ядерной ламиной.
В настоящее время общепринятым считается модульное строение ядерных каналов. Такая модель предусматривает структуру поры, состоящую из нескольких кольцевидных образований.
Внутри ядерной поры постоянно находится плотное вещество. Ее происхождение точно не известно, однако считается, что оно является одним из элементов ЯПК, за счет которого осуществляется транспортировка молекул от цитоплазмы к ядру и наоборот. Благодаря исследованию с использованием электронных микроскопов с высоким разрешением удалось выяснить, что плотная среда внутри ядерного канала способна менять свое месторасположение. Ввиду этого, считается, плотная внутренняя среда ЯПК является карго-рецепторным комплексом.
Транспортные функции ядерной оболочки возможны благодаря наличию ядерных поровых комплексов.
Виды ядерного транспорта
Транспортировка веществ через ядерную оболочку называется ядерно-цитоплазматическим транспортом веществ. Данный процесс предусматривает своеобразный обмен молекулами, синтезируемыми в ядре, и веществами, обеспечивающими жизнедеятельность самого ядра, импортируемыми из цитоплазмы.

Существуют такие виды транспортировки:
- Пассивная. Посредством данного процесса осуществляется перемещение небольших молекул. В частности, через пассивный транспорт происходит передача мононуклеотидов, минеральных компонентов, продуктов метаболического обмена. Процесс называется пассивным, так как протекает путем диффузии. Скорость прохождения через ядерную пору зависит от размера вещества. Чем оно меньше, тем выше скорость транспортировки.
- Активная. Предусматривает перенос через каналы внутри ядерной оболочки крупных молекул или их соединений. При этом, соединения не распадаются на мелкие частицы, что позволило бы увеличить скорость транспортировки. Данный процесс обеспечивает поступление в цитоплазму синтезируемых в ядре рибонуклеиновых молекул. Из внешнего цитоплазматического пространства за счет активного транспорта происходит перенос белков, необходимых для метаболических процессов.
Выделяют пассивный и активный транспорт белков, отличающийся механизмом действия.
Импорт и экспорт белков
Рассматривая функции ядерной оболочки, необходимо напомнить о том, что транспортировка веществ осуществляется в двух направлениях - из цитоплазмы в ядро и наоборот.
Импорт белковых соединений через мембраны к ядру осуществляется за счет наличия особых рецепторов, называемых транспортинами. Эти компоненты содержат запрограммированный сигнал, за счет которого происходит движение в необходимом направлении. и соединения, не обладающие таким сигналом, способны присоединятся к веществам, у которых он есть, и таким образом беспрепятственно перемещаться.
Важно отметить, что сигналы ядерного импорта обеспечивают избирательность поступления веществ в ядро. Многие образования, в числе которых полимеразы ДНК и РНК, а также белки, участвующие в процессах регуляции, не достигают ядра. Таким образом, ядерные поры представляют собой не только механизм транспортировки веществ, но и их своеобразной сортировки.
![]()
Сигнальные белки отличаются друг от друга. Ввиду этого, существует разница между скоростью перемещения через поры. Также они выполняют функцию источника энергии, так как для перемещения крупных молекул, транспортировка которых не возможна диффузным путем, необходимы дополнительные энергетические затраты.
Первый этап импорта белков заключается в присоединении к импортину (транспортину, обеспечивающему перенос через канал к ядру). Полученное в результате слияния сложное образование проходит через ядерную пору. После этого, с ним связывается другое вещество, за счет которого транспортируемый белок освобождается, а импортин возвращается обратно в цитоплазму. Таким образом, импорт в ядро представляет собой цикличный замкнутый процесс.
Транспорт веществ из ядра через оболочку в цитоплазматическое пространство осуществляется аналогичным образом. Исключением является то, что за перенос вещества-груза отвечают уже сигнальные белки, называемые экспортинами.
На первой стадии процесса белок (в большинстве случаев это молекулы РНК) связываются с экспортином и веществом, отвечающим за высвобождение транспортируемого субстрата. После перехода сквозь оболочку нуклеотид расщепляется, за счет чего переносимый белок высвобождается.
В целом, перенос веществ между ядром и цитоплазмой представляет собой цикличный процесс, осуществляемый за счет белков-транспортинов и веществ, отвечающих за высвобождение груза.
Ядерная оболочка при делении
Большинство клеток-эукариотов размножаются путем непрямого деления, которое называется митозом. Данный процесс предусматривает разделение ядра и других клеточных структур с сохранением одинакового количества хромосом. За счет этого сохраняется генетическая идентичность, полученная в результате деления клеток.

В процессе деления нуклеолемма выполняет еще одну важную функцию. После того как происходит разрушение ядра, внутренняя мембрана не позволяет хромосомам расходится на большие расстояния друг от друга. Хромосомы фиксируются на поверхности мембраны до момента полноценного деления ядер и формирования новой нуклеолеммы.
Ядерная оболочка, несомненно, принимает активное участие в клеточном делении. Процесс состоит из двух последовательных этапов - разрушения и перестройки.
Распад ядерной оболочки происходит в прометафазе. Разрушение мембран происходит стремительно. После распада хромосомы характеризуются хаотичным расположением в области ранее существовавшего ядра. В дальнейшем образуется веретено деления - биполярная структура, между полюсами которой формируются микротрубочки. Веретено обеспечивает деление хромосом и их распределение между двумя дочерними клетками.
Перераспределение хромосом и формирование новых ядерных мембран происходит в период телофазы. Точный механизм восстановления оболочек не известен. Распространенной является теория о том, что слияние частиц разрушенной оболочки происходит под действием везикул - мелких клеточных органелл, функция которых заключается в сборе и хранении питательных веществ.
Также образование новых ядерных мембран связывают с переформировкой эндоплазматического ретикулума. Из разрушенного ЭПР высвобождаются белковые соединения, которые постепенно обволакивают пространство вокруг нового ядра, в результате чего в дальнейшем образуется целостная мембранная поверхность.
Таким образом, нуклеолемма принимает непосредственное участие в процессе деления клетки путем митоза.
Ядерная оболочка - сложный структурный компонент клетки, выполняющий барьерные, защитные, транспортные функции. Полноценное функционирование нуклеолеммы обеспечивается , взаимодействием с другими клеточными компонентами и биохимическими процессами, протекающими в них.
Ядерная оболочка отграничивает содержимое ядра от цитоплазмы.
Она представляет собой систему из двух мембран, сливающихся между собой в зонах ядерных пор и отделяющихся содержимым перинуклеарного пространства (цистерна).
При световой микроскопии ядерная оболочка имеет вид довольно толстой, темноокрашенной линии, что обусловлено хроматином, прикрепляющимся к ее внутренней поверхности. Если рибонуклеиновые кислоты разрушить, то ядерная оболочка перестанет быть видимой при светооптических методах исследования.
Ядерная пора имеет общий диаметр около 20 нм, а канал ядерной поры - 9 нм. Это отверстия в ядерной оболочке, которые «полузакрыты» специальными белками (белками порового комплекса). В силу того что белки не совсем плотно прилежат друг к другу, между ними остается просвет, через который пассивно диффундируют по градиенту концентрации молекулы воды, растворенные в ней газы, неорганические ионы, низкомолекулярные органические вещества. Тем не менее, ядерная пора обеспечивает избирательный транспорт макромолекул. В результате содержимое ядра, значительно отличаясь от цитоплазмы по составу органических веществ высокого молекулярного веса (ферменты, макромолекулярные соединения), по составу низкомолекулярных веществ приближено к матриксу цитоплазмы.
Ядерная пора образована восемью периферическими и одной центральной белковыми гранулами. Центральная гранула связана с периферическими фибриллярными структурами. Иногда центральную гранулу рассматривают как трубочку с открытым каналом или как субъединицу рибосомы, транспортирующуюся через пору.
На внутренней и наружной поверхностях комплекса ядерной поры имеются высокоспецифичные рецепторы, обеспечивающие транспорт из ядра субъединиц рибосом, иРНК, тРНК и некоторых других веществ. В ядро избирательно транспортируются ферменты, ламины, гистоны. Транспорт макромолекул происходит активно, то есть белки комплекса ядерной поры обладают разной активностью (способны разрушать АТФ с высвобождением энергии). Частично такой транспорт происходит при расширении диаметра ядерной поры, например транспорт субъединиц рибосом.
У большинства клеток ядерные поры довольно многочисленны и располагаются на расстоянии 100…200 нм друг от друга, но при снижении белкового синтеза их число может уменьшаться. Это происходит при старении клетки и кариопикнозе.
Наружная ядерная мембрана по набору рецепторов и по составу аналогична гранулярной или гладкой ЭПС. Наружная ядерная мембрана участвует в синтезе полипептидных цепочек, которые поступают в перинуклеарное пространство, часто она связана с рибосомами и может сливаться с цистернами и каналами эндоплазматической сети.
Внутренняя ядерная мембрана участвует в формировании ядерной пластинки. Ядерная пластинка имеет толщину 80…300 нм, высокую электронную плотность, структурирует ядерную оболочку и перинуклеарный хроматин. Содержит белки - А-, В-, С-ламины, которые прикрепляются к белкам внутренней мембраны, выполняющим опорно-каркасные функции. Ламины образуют фибриллярные структуры, по функции близкие к промежуточным филаментам цитоплазмы. А-, В-, С-ламины в виде цепочки присоединены друг к другу. Кроме того, В-ламин связан и с интегральным белком внутренней мембраны кариолеммы. К ламинам прикрепляется хроматин (пристеночный), который может сильно конденсироваться.
Перинуклеарное пространство (цистерна) характеризуется низкой электронной плотностью. По химическому составу перинуклеарное пространство аналогично межмембранному пространству эндоплазматической сети, имеет толщину 200…300 нм и может расширяться в зависимости от функционального состояния клетки.
В разные периоды митотического цикла и при неодинаковой функциональной активности клеток в ядре выделяют некоторые особенности строения. В клетках с высокой скоростью анаболизма (синтеза) белков ядра обычно имеют большие размеры. В этом случае ядерная оболочка формирует инвагинации (впячивания), увеличивающие ее поверхность; содержит многочисленные ядерные поры; нарастает число ядрышек или они сливаются в одно-два крупных ядрышка с малой степенью конденсации. Нередко ядрышки смещаются к периферии ядра (ближе к ядерной оболочке). У малодифференцированных клеток преобладает эухроматин, диффузно распределенный в кариоплазме.
В зрелой клетке увеличивается объем гетерохроматина. По мере старения клетки и/или снижения ее синтетической активности содержание гетерохроматина увеличивается, ядрышки уплотняются и уменьшаются в размерах. В некоторых зрелых клетках сморщивание ядра сопровождается его сегментацией. В последующем такое сегментированное ядро может распадаться на крупные глыбки. Ядрышки и ядерные поры при этом подвергаются инволюции. Считывание генетической информации в этом случае практически прекращается. Такие процессы называют кариопикнозом - сморщивание ядра и кариорексисом - разрушение, распад ядра.
При патологических процессах, а также во время деления кариолемма может распадаться на пузырьки, содержимое ядра сливается с содержимым цитоплазмы. Данные изменения ядра называются кариолизисом. Кариолизису в патологических условиях может предшествовать набухание ядра с расширением перинуклеарного пространства (вакуолизация ядра).
У некоторых клеток усиление активности сопровождается не только увеличением синтетической активности, но и выраженным увеличением самой клетки - гипертрофией. В этом случае диплоидного набора хромосом в клетке оказывается недостаточно для поддержания ее жизнедеятельности и происходит полимеризация генетического материала с полиплоидизацией ДНК. В результате размеры ядра значительно увеличиваются, наблюдают одно-два крупных ядрышка, происходит их структурная перестройка.
Ядро есть в любой эукариотической клетке. Ядро может быть одно, или в клетке могут быть несколько ядер (в зависимости от ее активности и функции).
Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Ядерная оболочка состоит из двух мембран, разделенных перинуклеарным (околоядерным) пространством, между которыми находится жидкость. Основные функции ядерной оболочки: обособление генетического материала (хромосом) от цитоплазмы, а также регуляция двусторонних взаимоотношений между ядром и цитоплазмой.
Ядерная оболочка пронизана порами, которые имеют диаметр около 90 нм. Область поры (поровый комплекс) имеет сложное строение (это указывает на сложность механизма регуляции взаимоотношений между ядром и цитоплазмой). Количество пор зависит от функциональной активности клетки: чем она выше, тем больше пор (в незрелых клетках пор больше).
Основа ядерного сока (матрикса, нуклеоплазмы) – это белки. Сок образует внутреннюю среду ядра, играет важную роль в работе генетического материала клеток. Белки: нитчатые или фибриллярные (опорная функция), гетероядерные РНК (продукты первичной транскрипции генетической информации) и мРНК (результат процессинга).
Ядрышко – это структура, где происходят образование и созревание рибосомальных РНК (р-РНК). Гены р-РНК занимают определенные участки нескольких хромосом (у человека это 13–15 и 21–22 пары), где формируются ядрышковые организаторы, в области которых и образуются сами ядрышки. В метафазных хромосомах эти участки называются вторичными перетяжками и имеют вид сужений. Электронная микроскопия выявила нитчатый и зернистый компоненты ядрышек. Нитчатый (фибриллярный) – это комплекс белков и гигантских молекул-предшественниц р-РНК, которые дают в последующем более мелкие молекулы зрелых р-РНК. При созревании фибриллы превращаются в рибонуклеопротеиновые гранулы (зернистый компонент).
Хроматин получил свое название за способность хорошо прокрашиваться основными красителями; в виде глыбок он рассеян в нуклеоплазме ядра и является интерфазной формой существования хромосом.
Хроматин состоит в основном из нитей ДНК (40 % массы хромосомы) и белков (около 60 %), которые вместе образуют нуклеопротеидный комплекс. Выделяют гистоновые (пять классов) и негистоновые белки.
Гистонам (40 %) принадлежат регуляторная (прочно соединены с ДНК и препятствуют считыванию с нее информации) и структурная функции (организация пространственной структуры молекулы ДНК). Негистоновые белки (более 100 фракций, 20 % массы хромосомы): ферменты синтеза и процессинга РНК, репарации редупликации ДНК, структурная и регуляторная функции. Кроме этого, в составе хромосом обнаружены РНК, жиры, полисахариды, молекулы металлов.
В зависимости от состояния хроматина выделяют эухромати-новые и гетерохроматиновые участки хромосом. Эухроматин отличается меньшей плотностью, и с него можно производить считывание генетической информации. Гетерохроматин более компактен, и в его пределах информация не считывается. Выделяют конститутивный (структурный) и факультативный гетерохро-матин.
5. Строение и функции полуавтономных структур клетки: митохондрий и пластид
Митохондрии (от гр. mitos – «нить», chondrion – «зернышко, крупинка») – это постоянные мембранные органеллы округлой или палочковидной (нередко ветвящейся) формы. Толщин – 0,5 мкм, длина – 5–7 мкм. Количество митохондрий в большинстве животных клеток – 150-1500; в женских яйцеклетках – до нескольких сотен тысяч, в сперматозоидах – одна спиральная митохондрия, закрученная вокруг осевой части жгутика.
Основные функции митохондрий:
1) играют роль энергетических станций клеткок. В них протекают процессы окислительного фосфорилирования (ферментативного окисления различных веществ с последующим накоплением энергии в виде молекул аденозинтрифосфата – АТФ);
2) хранят наследственный материал в виде митохондриаль-ной ДНК. Митохондрии для своей работы нуждаются в белках, закодированных в генах ядерной ДНК, так как собственная митохондриальная ДНК может обеспечить митохондрии лишь несколькими белками.
Побочные функции – участие в синтезе стероидных гормонов, некоторых аминокислот (например, глютаминовой). Строение митохондрий
Митохондрия имеет две мембраны: наружную (гладкую) и внутреннюю (образующую выросты – листовидные (кристы) и трубчатые (тубулы)). Мембраны различаются по химическому составу, набору ферментов и функциям.
У митохондрий внутренним содержимым является матрике – коллоидное вещество, в котором с помощью электронного микроскопа были обнаружены зерна диаметром 20–30 нм (они накапливают ионы кальция и магния, запасы питательных веществ, например, гликогена).
В матриксе размещается аппарат биосинтеза белка органеллы: 2–6 копий кольцевой ДНК, лишенной гистоновых белков (как у прокариот), рибосомы, набор т-РНК, ферменты редупликации, транскрипции, трансляции наследственной информации. Этот аппарат в целом очень похож на таковой у прокариот (по количеству, структуре и размерам рибосом, организации собственного наследственного аппарата и др.), что служит подтверждением симбиоти-ческой концепции происхождения эукариотической клетки.
В осуществлении энергетической функции митохондрий активно участвуют как матрикс, так и поверхность внутренней мембраны, на которой расположена цепь переноса электронов (цитохро-мы) и АТФ-синтаза, катализирующая сопряженное с окислением фосфорилирование АДФ, что превращает его в АТФ.
Митохондрии размножаются путем перешнуровки, поэтому при делении клеток они более или менее равномерно распределяются между дочерними клетками. Так, между митохондриями клеток последовательных генераций осуществляется преемственность.
Таким образом, митохондриям свойственна относительная автономность внутри клетки (в отличие от других органоидов). Они возникают при делении материнских митохондрий, обладают собственной ДНК, которая отличается от ядерной системой синтеза белка и аккумулирования энергии.
Пластиды
Это полуавтономные структуры (могут существовать относительно автономно от ядерной ДНК клетки), которые присутствуют в растительных клетках. Они образуются из пропластид, которые имеются у зародыша растения. Отграничены двумя мембранами.
Выделяют три группы пластид:
1) лейкопласты. Имеют округлую форму, не окрашены и содержат питательные вещества (крахмал);
2) хромопласты. Содержат молекулы красящих веществ и присутствуют в клетках окрашенных органов растений (плодах вишни, абрикоса, помидоров);
3) хлоропласты. Это пластиды зеленых частей растения (листьев, стеблей). По строению они во многом схожи с митохондриями животных клеток. Наружная мембрана гладкая, внутренняя имеет выросты – ламелосомы, которые заканчиваются утолщениями – тилакоидами, содержащие хлорофилл. В строме (жидкой части хлоропласта) содержатся кольцевая молекула ДНК, рибосомы, запасные питательные вещества (зерна крахмала, капли жира).







