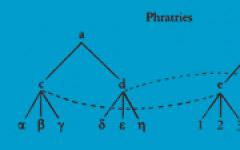
Регуляция репродуктивной функции определяется взаимодействием с нейроэндокринной системой, включающей в себя центральные (интегри-рующие) отделы, промежуточные звенья и периферические, эффекторные структуры. Взаимосвязь желез внутренней секреции, входящих в систему регуляции репродуктивной функции, и органов—мишеней определяется прежде всего специфическими рецепторами. Рецепторы располагаются на цитоплазматической мембране и затем поступают в ядро клетки, связываясь с ДНК. Ядро гормонально зависимой клетки является акцептором не только гормонов, но и аминопептидов, инсулина, глюкагона и др. ДНК после связи с гормонами оказывает специфическое действие на метаболизм.
В регуляции репродуктивной системы выделяют пять уровней, которые действуют по принципу прямой и обратной связи благодаря наличию во всех звеньях цепи рецепторов к половым и гонадотропным гормонам.
Первым (высшим) уровнем регуляции репродуктивной системы являются кора головного мозга, гипоталамус и экстрагипоталамические церебраль-ные структуры, лимбическая система, гиппокамп, миндалевидное тело.
О роли ЦНС в регуляции менструального цикла было известно до выде-ления гормонов и нейросекретов. Наблюдали прекращение менструаций при стрессах, при очень большом желании иметь беременность или при боязни забеременеть у женщин с неустойчивой психикой. В настоящее время в коре головного мозга, в гипоталамусе и экстрагипоталамических структурах выявлены специфические рецепторы к половым гормонам. Кроме того, в ответ на внешние и внутренние раздражители в коре и под-корковых структурах происходят синтез и выделение нейротрансмиттеров и нейропептидов, которые влияют прежде всего на гипоталамус, способс-твуя синтезу и выделению рилизинг-гормона.
Корой головного мозга выделяются эндогенные опиодные пептиды (ЭОП): энкефалины, эндорфины и динорфины. Эти субстанции обнару-живаются не только в различных структурах мозга и вегетативной нервной системы, но и в печени, легких, поджелудочной железе и других органах, а также в некоторых биологических жидкостях (плазма крови, содержимое фолликула). По современным представлениям, ЭОП оказывает воздейс-твие на гипоталамус.
К важнейшим нейротрансмиттврам, т.е. веществам-передатчикам, отно-сятся норадреналин, дофамин, гамма-аминомасляная кислота (ГАМК), ацетилхолин, серотонин и мелатонин.
Церебральные нейротрансмиттеры регулируют выработку гонадотропин-рилизинг гормона (ГнРГ): норадреналин, ацетилхолин и ГАМК сти-мулируют их выброс, а дофамин и серотонин оказывают противоположное действие. Нейропептиды (эндогенные опоидные пептиды, кортикотропин-рилизинг фактор и галанин) также влияют на функцию гипоталамуса и на сбалансированность функционирования всех звеньев репродуктивной системы.
Вторым уровнем регуляции репродуктивной системы является гипота-ламус, в котором секретируются стимулирующие (либерины) и блокирую-щие (статины) нейрогормоны. Клетки, которые выделяют нейрогормоны (пептидергические нейронные), обладают свойствами как нейронов, так и эндокринных желез.
Гипоталамус секретирует ГнРГ, содержащие фолликулостимулирующий (РГФСГ — фоллиберин) и лютеинизирующий (РГЛГ — люлиберин) гормоны, которые воздействуют на гипофиз.
Рилизинг-гормон ЛГ (РГЛГ — люлиберин) выделен, синтезирован и подробно описан. Выделить и синтезировать рилизинг-фолликулостимулирующий гормон до настоящего времени не удалось. Однако установлено, что декапептид РГЛГ и его синтетические аналоги стимулируют выделение гонадотрофами не только ЛГ, но и ФСГ. В связи с этим принят один термин для гонадоторопных либеринов — гонадотропин-рилизинг гормон (ГнРГ), по сути являющийся синонимом РГЛГ.
Секреция ГнРГ генетически запрограммирована и носит пульсирующий (цирхоральный) характер: пики усиленной секреции гормона продолжитель-ностью несколько минут сменяются 1—3-часовыми интервалами относитель-но низкой секреторной активности. Частота и амплитуда секреции ГнРГ в преовуляторный период на фоне максимального выделения эстрад иола зна-чительно больше, чем в раннюю фолликулярную и лютеиновую фазы.
Деятельность гипоталамуса тесно связана с функцией гипофиза.
К третьему уровню регуляции относится передняя доля гипофиза (аденогипофиз), в котором синтезируются фолликулостимулирующий гор-мон, или фоллитропин (ФСГ); лютеинезирующий, или лютропин (ЛГ); пролактин (ПрЛ); адренокортикотропный (АКТГ); соматотропный (СТГ); тиреотропный или тиролиберин (ТТГ); ФСГ, ЛГ, ПрЛ воздействуют на яичник. ПрЛ стимулирует рост молочных желез и лактацию, контролирует секрецию прогестерона желтым телом путем активации образования в них рецепторов к Л Г.
Синтез ПрЛ аденогипофизом находится под тоническим блокирующим контролем дофамина, или пролактинингибирующего фактора. Ингибиция синтеза ПрЛ прекращается во время беременности, лактации. Основным стимулятором синтеза ПрЛ является ТТГ, синтезируемый в гипоталамусе.
Остальные гормоны гипофиза влияют на соответствующие их названию железы внутренней секреции. Только при сбалансированном выделении каждого из гомонов гипофиза возможна нормальная функция.репродук-тивной системы.
К четвертому уровню регуляции репродуктивной функции относятся периферические эндокринные органы (яичники, надпочечники, щито-видная железа). Основная роль принадлежит яичникам, а другие железы выполняют собственные специфические функции, одновременно подде-рживая нормальное функционирование репродуктивной системы.
Пятый уровень регуляции репродуктивной функции составляют чувс-твительные к колебаниям уровней половых стероидов внутренние и вне-шние отделы репродуктивной системы (матка, маточные трубы, слизистая оболочка влагалища), а также молочные железы. Наиболее выраженные циклические изменения происходят в эндометрии.
Цикличность системы, регулирующей репродуктивную функцию, определяется прямой и обратной связью между отдельными звеньями. Так, ФСГ, благодаря рецепторам в фолликулярных клетках яичника, стиму-лирует выработку эстрогенов (прямая связь). Эстрогены, накапливаясь в большом количестве, блокируют выработку ФСГ (обратная связь).
Во взаимодействии звеньев репродуктивной системы различают «длин-ную», «короткую» и «ультракороткую» петли. «Длинная» петля — воздействие через рецепторы гипоталамо-гипофизарной системы на выработку половых гормонов. «Короткая» петля определяет связь между гипофизом и гипоталаму-сом, «ультракороткая» петля — связь между гипоталамусом и нервными клет-ками, которые осуществляют локальную регуляцию с помощью нейротранс-миттеров, нейропептидов, нейромодуляторов и электрических стимулов.
О деятельности репродуктивной системы во время менструального цикла судят по тестам функциональной диагностики: базальной темпера-туре, симптому зрачка, симптому расширения шеечной слизи, морфологи-ческому состоянию эндометрия.
Базальную температуру измеряют в прямой кишке ежедневно утром до вставания с постели. Результаты измерения наносят на график, по которо-му можно судить о фазности яичниковых изменений. При нормальном овуляторном двухфазном цикле в первую фазу температура ниже 37 °С. Перед овуляцией она несколько снижается, а в период овуляции повы-шается до 37,6—37,8 °С, а затем в лютеиновую фазу остается субфебрильной. .Перед менструацией температура вновь снижается. При наступлении беременности температура в прямой кишке под влиянием прогестерона остается субфебрильной.
Симптом зрачка отражает секрецию слизи железами шейки матки под влиянием эстрогенов. В предовуляторные дни секреция слизи увеличива-ется, наружное отверстие шеечного канала приоткрывается и при осмотре в зеркалах напоминает зрачок. В соответствии с диаметром видимой в шейке слизи выраженность симптома зрачка определяют плюсами. В период ову-ляции симптом зрачка составляет +++, затем под влиянием прогестерона ко дню менструального цикла он равен +, а затем исчезает.
Симптом растяжения шеечной слизи связан с ее изменениями под влия-нием эстрогенов. Растяжимость слизи определяют с помощью корнцанга. Берут каплю слизи из шеечного канала и, раздвигая бранши, смотрят, на сколько миллиметров растягивается слизь. Максимальное растяжение нити (10—12 мм) происходит в период наибольшей концентрации эстроге-нов, соответствующий овуляции.
Кариопикнотический индекс (КПИ). Под влиянием эстрогенов в сли-зистой оболочке влагалища, особенно в его верхней трети, происходит ороговение поверхностных клеток, ядра которых становятся пикнотическими. В мазке, взятом из заднего свода влагалища, при микроскопическом исследовании определяют соотношение поверхностных, ороговевающих и промежуточных клеток. Чем больше процент ороговевающих клеток, тем больше в организме эстрогенов. Максимальное число ороговевающих кле-ток выявляется в предовуляторный период (80—88%), в первые дни после менструации — 20—40%, перед менструацией — 20—25%. Двухфазность изменений во время менструального цикла можно определить при гисто-логическом исследовании эндометрия. В 1-й день менструации при овуляции и образовании желтого тела в слизистой оболочке матки (цуг-биопсия) видны изменения, соответствующие фазе секреции.
Правильнее говорить не о менструальном цикле, а о репродуктивной системе, которая подобно другим является функциональной системой (по Анохину, 1931).
Функциональная система - это интегральное образование включающее центральные и периферические звенья и работающие по принципу обратной связи, то есть с обратной афферентацией по конечному эффекту.
Все другие системы поддерживают гомеостаз, а репродуктивная система поддерживает воспроизводство - существование человеческого рода.
Функциональной активности система достигает к 16-17 годам. К 40 годам репродуктивная функция угасает, а к 50 годам угасает гормональная функция.
Репродуктивная система организована по иерархическому принципу. В ней выделяют 5 уровней, каждый из которых регулируется вышележащими структурами по механизму обратной связи.
ПЕРВЫЙ УРОВЕНЬ: ткани мишени - это точки приложения действия половых гормонов: половые органы: матка, трубы, шейка матки, влагалище, молочные железы, волосяные фолликулы, кожа, кости, жировая ткань. Цитоплазма этих клеток содержит строго специфические рецепторы к половым гормонами: эстрадиолу, прогестерону, тестостерону. Эти рецепторы есть в нервной системе.
К первому уровню кроме цАМФ относятся и простогландины. Из всех этих органов мишеней наибольшие изменения как в первом уровне регуляции происходят в матке.
В матке под действие половых гормонов в первую фазу цикла (фолликулиновую происходят процессы пролиферации, идет рост и размножение желез слизистой матки, они достигают своего развития к 14 дню цикла, когда в яичниках происходит овуляция и затем под действием выделяющегося прогестерона в матке происходит секреторные преобразования. Только секреторно преобразованная слизистая способна воспринять оплодотворенное яйцо. Если же оплодотворение не происходит, то в ответ на падение уровня гормонов выделенных яичниками в матке происходят еще 2 фазы:
" отторжение слизистой полости матки – десквамация
" регенерация идущая параллельно с десквамацией
ВТОРОЙ УРОВЕНЬ - ЯИЧНИКИ. В клетках теки и гранулезы яичника имеются рецепторы к гонадотропным гормонам. У приматов и у человека на протяжении цикла развивается один фолликул - лидирующий или доминантный - определяет дальше развитие изменений происходящих не только в яичнике, но и во всей гипоталамо-гипофизарной системы. В начале цикла доминантный фолликул до 2 мм, когда он достигает конечного развития он бывает до 21 мм, а по некоторым данным и больше.
Подъем уровня эстрогенов стимулирует выделение лютенизирующего гормона и в яичниках происходит разрыв этого фолликула и яйцеклетка выделяется в брюшную полость, а на месте лопнувшего фолликула образуется желтое тело. Желтое тело начинает выделять в основном прогестерон, эстрогены и андрогены. В регрессе желтого тела, если не происходит оплодотворения участвуют простогландины.
Все стероидный гормоны образуются из холестерина, в синтез участвуют гонадотропные гормоны: ФСГ и ЛГ и ароматазы под влиянием которых из андрогенов образуются эстрогены.
ТРЕТИЙ УРОВЕНЬ - ПЕРЕДНЯЯ ДОЛЯ ГИПОФИЗА (ФСГ, ЛГ, ПРОЛАКТИН). ФСГ стимулирует в яичнике рост фолликула, пролиферацию гранулезных клеток, вместе с ЛГ стимулирует выделение эстрогенов, увеличивает содержание ароматаз.
Повышение выделения ЛГ при созревшем доминантном фолликуле вызывает овуляцию. Затем ЛГ стимулирует выделение прогестерона желтым телом. Рассвет желтого тела определяется дополнительным влиянием пролактина.
Основная биологическая функция пролактина - рост молочных желез и регуляция лактации. Кроме этого он обладает жиромобилизующим эффектом и понижает АД. Повышение в организм пролактина ведет к нарушению менструального цикла.
ЧЕТВЕРТЫЙ УРОВЕНЬ - ГИПОФИЗОТРОПНАЯ ЗОНА ГИПОТАЛАМУСА.
Состоит из 3-х ядер: вентро и дорсомедиальных, аркуатных ядер. В них образуются рилизинг-факторы, которые стимулируют выделение тропных гормонов.
РФ-ЛГ выделен и синтезирован, РФ-ФСГ не выделен, поэтому в литературе пользуются терминов РГ-ЛГ.
Долгое время считали что у человека, как и у грызунов имеется 2 центра секреции РГ-ЛГ: циклический и тонический. Но это мнение было опровергнуто в 70 гг. и было доказано что у приматов и у человека единственным место, где образуется РГ-ЛГ являются аркуатные ядра. Секреция РГ-ЛГ генетически запрограммирована и происходит в определенном пульсирующем режиме с частотой 1 раз в час. Этот ритм называется цирхаральным (часовой).
Область аркуатных ядер гипоталамуса называется аркуатным асцилятором. Цирхаральный ритм подтвержден путем прямого измерения ЛГ в портальной системе ножки гипофиза и яремной вены у женщин с нормальной функцией. Эти исследования позволили обосновать гипотезу о пермессивной (запускающей) роли РГ-ЛГ в функции репродуктивной системы.
Цирхаральный режим выделения РГ-ЛГ формируется в пубертатном возрасте и является показателем зрелости нейроструктур гипоталамуса. Определенная роль в регуляции выделения РГ-ЛГ принадлежит эстрадиолу. В преовуляторном периоде на фоне максимального уровня эстрадиола в крови величина выброса РГ-ЛГ значительно выше в раннюю фоликулиновую и лютеиновую фазу. Однако различий в частоте выброса нет. Есть сообщение о наличие пролактин-РГ, однако он не выделен, но доказано что тиролиберин стимулирует выделение пролактина. Основная роль в выделении пролактина принадлежит фундикулярной области гипоталамуса. Дофамин тормозит выделение пролактина. Антагонисты дофамин - резерпин, аминазин вызывают усиление выброса пролактина. Поэтому после родов, если у женщины сниженная лактация рекомендуется применять резерпин.
Экстрагипоталамические центральные структуры, которые воспринимают импульсы из внешней Среды и интерорецепторов организма и передают их через нейротрансмиттеры. Последние годы выделены и синтезированы классические синаптические нейротрансмиттеры (биогенные амины: норадреналин, серотонин, морфиноподобные соединения - эндорфины и энкефалины, последние составляют 90%). Энкефалины обнаружены во всех структурах головного мозга. Они подавляют секрецию ЛГ, поэтому наркоманы, как правило, страдают бесплодием. А их антагонисты резко усиливают выброс РГ-ЛГ. Изучение влияние коры головного мозга сложно. Центры регуляции в коре не обнаружены, но клиника нарушений менструального цикла подтверждает эти влияния.
В регуляции функции репродуктивной системы основными являются:
" пульсирующая цирхаральная секреция РГ-ЛГ нейронов гипоталамуса
" регуляция выделения ЛГ и ФСГ эстрадиолом по типу положительной и отрицательной связи
Нормальный менструальный цикл является результатом нейрогормональных взаимоотношений между ЦНС, гипофизом, яичниками и маткой. Нарушения в любой из этих звеньев может привести к нарушению менструального цикла.
Наиболее частыми причинами нарушение менструальной функции могут быть:
1. Тяжелые интоксикации
2. Инфекционные заболевания типа сепсиса, туберкулеза, паротита, гриппа и т.д.
3. Алиментарная дистрофия
4. Авитаминозы
5. Ожирение
6. Профессиональные вредности
7. Заболевания других систем и органов: печени, почек, сердца и крови.
8. Психоневрологические заболевания: ушибы головного мозга, контузии, опухоли ЦНС.
9. Эндокринные заболевания: сахарный диабет, заболевания надпочечников, щитовидной железы, гипофиза, синдром Шихана, кахексия, синдром Симмондса, аденома гипофиза.
10. Ионизирующая радиация
11. Врожденные, генетические заболевания
12. Воспалительные из невоспалительные заболевания самой половой сферы (придатков и матки)
Таким образом, все может вызвать нарушения менструальной функции.
В звеньях патогенеза следует различать:
1. Первичное поражение гипоталамо-гипофизарной системы, которое в конечном итоге приведет к нарушению функции яичников
2. Первичное заболевание самих яичников и матки
Действие повреждающих факторов непосредственно на фолликулярный аппарат яичников может привести к нарушению их функции разной степени выраженности: от функциональных нарушений до аменореи.
Классификация основных нарушений репродуктивной системы в принципе сложна. Их в литературе вы встретите не одну, не две. Но, несмотря на всю сложность строения репродуктивной системы, основной формой ее нарушения является нарушения основного момента, который характерен для нормального менструального цикла в яичниках, то есть нарушения того для чего эта система существует - нарушение овуляции.
При нарушениях наблюдаются ановуляция, то есть отсутствие овуляции, а клинически могут наблюдаться незначительные нарушения менструальной функции или только одно бесплодие, а может наблюдаться и при кровотечениях и при присутствии месячных. Учитывая всю эту сложность мы даем саму упрощенную классификацию, хотя эта классификация в итоге по конечному нарушению самой менструальной функции, то есть по клиническому проявлению:
" гипоменструальный синдром
" гиперменструальный синдром
19. Центральный уровень регуляции репродуктивной функции
См. предыдущий вопрос…
20. Роль яичников в регуляции менструального цикла
См. вопрос №18…
21. Подовые гормоны и их биологические свойства
См. вопрос №18…
22. Физиотерапевтические методы лечения в гинекологии [-]
23. Санаторно - курортное лечение гинекологических больных [-]
*НЕЙРОЭНДОКРИННАЯРЕГУЛЯЦИЯ
РЕПРОДУКТИВНОЙ
ФУНКЦИИ
Доцент кафедры акушерства и
гинекологии
к.м.н. Карпова Ирина АдамовнаРЕПРОДУКЦИЯ - размножение,
воспроизведение себе подобных
Осуществляется за счет двух процессов:
подготовки организма женщины
- к оплодотворению яйцеклетки
- и к гестации (беременности)
Обе стадии вместе длятся в среднем 28дней
(от 21 до 33)
В этот период обеспечивается
-созревание яйцеклетки в яичниках
(овариальный цикл)
-и подготовка эндометрия в матке
(маточный цикл)ЦИКЛИЧЕСКИЙ ТИП ФУНКЦИОНИРОВАНИЯ
Является особенностью женской
репродуктивной системы, в отличие от
мужской репродуктивной системыМЕНСТРУАЛЬНЫЙ ЦИКЛ
Одно из самых ярких проявлений циклических
процессов происходящих в организме женщины.
Характеризуется циклическими изменениями во всех
звеньях репродуктивной системы, внешним
проявлением которых является менструация.МЕНСТРУАЦИЯ (от menstruus - месячный)
Периодически появляющиеся непродолжительные
маточные кровотечения, обусловленные
отторжением функционального слоя эндометрия в
конце двухфазного (овуляторного) менструального
цикла.Менструальная функция отражает
способность репродуктивной системы
женщины к деторождению
Первая менструация в жизни женщины менархе
(menarche) наступает в 12-13 ±1,5-2 года
У 60 % женщин средняя продолжительность
менструального цикла составляет 28 дней с
колебаниями от 21 до 35 дней.
Величина кровопотери в менструальные дни 40-60
мл (в ср. 50 мл)
Продолжительность нормальной менструации от 2 до
7 дней.
Последняя менструация (менопауза) - 50,8 лет.НОРМАЛЬНЫЙ МЕНСТРУАЛЬНЫЙ ЦИКЛ ВКЛЮЧАЕТ:
- циклические
изменения в системе гипоталамусгипофиз-яичники (овариальный или яичниковый
цикл, созревание и выход яйцеклетки)
-циклические изменения в эндометрии (маточный
цикл, пролиферация и секреторная трансформация
эндометрия)
- многообразные физиологические сдвиги в
различных функциях организма (менструальная
волна)
ОСУЩЕСТВЛЯЕТСЯ С УЧАСТИЕМ 5 ЗВЕНЬЕВ (УРОВНЕЙ)
Центральные
1. Кора головного мозга
2. Гипоталамус
3. Гипофиз
Периферические
4. Яичники
5. МаткаКОРА ГОЛОВНОГО МОЗГА
ВЕДУЩУЮ РОЛЬ ПОДТВЕРЖДАЮТ
Нарушения овуляции при различных острых и
хронических стрессах (потеря работы, семейные
конфликты, экзаменационная сессия).
Нарушение менструального цикла при перемене
климатогеографических зон, ритма работы.
Хорошо известно прекращение менструаций в условиях
военного времени (аменорея военного времени).
У психически неуравновешенных женщин, страстно
желающих иметь ребенка, менструации также могут
прекратиться.КОРА ГОЛОВНОГО МОЗГА
Точная
локализация центров регуляции функции
половой системы в коре головного мозга не
установлена.
ИНФОРМАЦИЯ ИЗ ВНЕШНЕЙ СРЕДЫ и
ИНТЕРОРЕЦЕПТОРОВ
По СПЕЦИАЛИЗИРОВАННЫМ НЕЙРОНАМ ГОЛОВНОГО
МОЗГА
Через систему НЕЙРОТРАНСМИТТЕРОВ или
НЕЙРОМЕДИАТОРОВ
Поступает в НЕЙРОСЕКРЕТОРНЫЕ ЯДРА ГИПОТАЛАМУСА
Синтез гипофизотропных РИЛИЗИНГ-ГОРМОНОВ
(ГТ- РГ - ГОНАДОЛИБЕРИН)
Отводится наиболее важная роль в регуляции
функции гипоталамо-гипофизарной системы
Норадреналин стимулирует овуляторный
выброс гонадолиберина
Дофамин поддерживает секрецию ГТ-РГ в
аркуатных ядрах гипоталамуса, тормозит
выделение аденогипофизом пролактина (ПИФ пролактинингибирующий фактор)
Серотонин контролирует (тормозит)
циклическую секрецию гонадолиберинаМОРФИНОПОДОБНЫЕ ОПИОИДНЫЕ
НЕЙРОПЕПТИДЫ
(ЭНДОРФИНЫ, ЭНКЕФАЛИНЫ, ДОНОРФИНЫ)
подавляют секрецию лютеинизирующего
гормона (ЛГ)
фолликулостимулирующего гормона (ФСГ)
тиреотропного гормона (ТТГ)
повышают секрецию пролактина (ПРЛ)
гормона роста (СТГ) Таким
образом, кора головного мозга
(ЦНС) осуществляет контроль над
гипоталамо-гипофизарной системой
посредством нейротрансмиттеров,
которые контролируют активность
нейроэндокринных нейронов
гипоталамуса.ГИПОТАЛАМУС
Важнейшее звено в регуляции
менструального цикла.
В гипоталамусе нервные сигналы
преобразуются в эндокринные.
Т.е. гипоталамус связывает нервную и
эндокринную системы.
Ядра гипоталамуса представляют собой
скопления нервных клеток, обладающих
нейросекреторной активностью.ГИПОТАЛАМУС
Два типа эндокринных нейронов гипоталамуса
1. Крупноклеточные нейроны супраоптического и
паравентрикулярного ядер синтезируют и
секретируют гормоны задней доли гипофиза -
окситоцин и антидиуретический гормон
(вазопрессин), которые депонируются в
нейрогипофизе (задней доле гипофиза).
2. Мелкоклеточные нейроны вентро- и
дорсомедиальных, аркуатных ядер вырабатывают
специфические нейросекреты (либерины и
статины).
Либерины или рилизинггормоны (рилизингфакторы)
(realizing factors):
1) кортиколиберин - стимулирует секрецию
АКТГ и МСГ (меланоцитостимулирующий
гормон);
2) соматолиберин - стимулирует секрецию СТГ;
3) тиреолиберин - стимулирует секрецию ТТГ и
пролактина;
4) гонадолиберин - стимулирует секрецию ЛГ и
ФСГ.ГИПОТАЛАМУС
Статины:
1) соматостатин - подавляет секрецию СТГ (в
меньшей степени -ТТГ);
2) дофамин - подавляет секрецию пролактина.ГИПОТАЛАМУС
Гонадолиберин синтезируется в аркуатных ядрах
медиобазального гипоталамуса, получивших название
«аркуатный осциллятор».
«Аркуатный осциллятор» - кольцевая структура,
объединяющую много сотен разных типов нейронов и
функционирующая, как замкнутая цепь.
«АО» выполняет роль генератора импульсного ритма
секреции ГТ-РГ.
Секреция ГТ-РГ аркуатными ядрами генетически
запрограммирована и происходит в пульсирующем, или
цирхоральном режиме (цирхоральный - часовой).
Самая сложная по строению и функции эндокринная
железа, место синтеза всех тропных гормонов.
В аденогипофизе синтезируются
Адренокортикотропный гормон (АКТГ) в кортикотрофах
Тиреотропный гормон (ТТГ) в тиреотрофах
Гормон роста (ГР) в соматотрофах
Пролактин (ПРЛ) в лактотрофах (ацидофильных
клетках аденогипофиза)
Фолликулостимулирующий (ФСГ) и лютеинизирующий
гормоны (ЛГ) синтезируются в клетках одного типа - в
гонадотрофах (базофильных клетках аденогипофиза)ГИПОФИЗ
В задней доле (нейрогипофиз)
накапливаются и выделяются окситоцин и
антидиуретический гормон (нейросекрет
гипоталамуса).ГИПОФИЗ
Часовые импульсы ГТ-РГ стимулируют гонадотрофы к
импульсам секреции ФСГ и ЛГ с интервалами между
отдельными импульсами 90–120 мин.
При этом частота импульсов меняется на протяжении
менструального цикла - от 1 импульса в 90 мин в начале
цикла с учащением до 1 импульса в 60 мин к моменту
овуляции.
А во второй фазе цикла импульсы происходят с интервалом в
3–4 ч.
Железой-мишенью для ФСГ и ЛГ является яичник.ГИПОФИЗ
ФСГ
рост, развитие и созревание фолликулов
- пролиферация и дифференцировка клеток
гранулезы
- увеличение содержания ароматаз в зреющем
фолликуле
- синтез ингибина
- синтез ЛГ-рецепторов в гранулезных клетках,
необходимых для продукции прогестерона в
лютеиновой фазе.ГИПОФИЗ
ЛГ
- стимулирует первые этапы синтеза андрогенов в
тека-клетках фолликула
- синтез Е2, овуляцию, синтез прогестерона в
лютеинизированных клетках гранулезы (в желтом
теле).
Совместно ФСГ и ЛГ
- стимулируют секрецию эстрогенов, овуляцию,
- лютеинизацию гранулезных клеток фолликула
- секрецию прогестерона в лютеинизированных
клетках гранулезы (в желтом теле).ДВА ВИДА СЕКРЕЦИИ ГОНАДОТРОПНЫХ
ГОРМОНОВ
Тонический тип характеризуется низким уровнем
частоты и амплитуды импульсов в течение всего цикла и
способствует развитию фолликулов и продукции ими
эстрогенов.
Циклический тип проявляется в резком возрастании
частоты и амплитуды импульсов (пики) выделения
гонадотропинов в определенные дни цикла (пик ФСГ - на
7-й и на 14-й день; пик ЛГ - на 14-й (овуляторный) и
22–23-й дни цикла), обеспечивая смену фаз низкой и
высокой секреции гормонов.СОДЕРЖАНИЕ ГОРМОНОВ В ПЛАЗМЕ КРОВИ В
ТЕЧЕНИЕ МЕНСТРУАЛЬНОГО ЦИКЛАПРОЛАКТИН
Синтез пролактина аденогипофизом находится
под тоническим блокирующим контролем
дофамина, или пролактинингибирующего
фактора.
Ингибиция прекращается во время беременности,
лактации.
Основным стимулятором синтеза пролактина
является тиролиберин
Секреция пролактина носит импульсный характер,
происходит в циркадианном ритме - с четкими
периодическими изменениями в течение суток
(уровень пролактина повышается в ночное время).ПРОЛАКТИН
Биологическое действие ПРЛ
- стимулирует рост молочных желез и регулирует
лактацию
- обладает жиромобилизующим и гипотензивным
эффектом
- повышает активность β-клеток поджелудочной
железы, способствуя - развитию инсулиновой
резистентности
- в повышенных количествах тормозит рост и
созревание фолликула
- физиологические дозы ПРЛ стимулируют функцию
желтого тела и выработку прогестерона, а также
тормозят синтез андрогенов в яичниках.РЕГУЛЯЦИЯ МЕНСТРУАЛЬНОЙ ФУНКЦИИ
Построена по иерархическому принципу - каждый из
уровней регулируется вышележащей структурой
нейрогуморальным путем
Реализуя механизм
- отрицательной обратной связи
(снижение уровня периферических гормонов стимулирует
синтез и выделение тропных гормонов гипофиза)
- положительной обратной связи, когда в ответ на
максимальное повышение уровня эстрадиола (Е2) в
преовуляторном периоде возрастают синтез и секреция
гонадотропинов
Ведущая роль принадлежит
гипоталамо-гипофизарной системе, которая определяет
состояние
всей эндокринной системы.
контролируется гонадолиберином,
выделяющимся в пульсирующем режиме.
Амплитуда импульсов гонадотропинов меняется в
зависимости от фазы менструального цикла и
определяется гормонами яичника посредством
механизма отрицательной и положительной
обратной связи через рецепторы, расположенные
в гипоталамусе и гипофизе (так называемая
длинная петля обратной связи).ЯИЧНИКИ
Выполняют в организме женщины эндокринную и
генеративную функции.
Во время менструального цикла в яичниках
происходит рост фолликулов и созревание
яйцеклетки, которая в результате становится
готовой к оплодотворению.
Одновременно в яичниках вырабатываются
половые гормоны, обеспечивающие изменения в
слизистой оболочке матки, способной воспринять
оплодотворенное яйцо.ЯИЧНИКИ
Циклические изменения в яичниках включают три
основных процесса:
1. Рост фолликулов и формирование доминантного
фолликула.
2. Овуляция.
3. Образование, развитие и регресс желтого тела.
При рождении девочки в яичнике находятся 2 млн.
фолликулов, 99 % которых подвергаются атрезии в
течение всей жизни.
Под процессом атрезии понимается обратное
развитие фолликулов на одной из стадий его
развития.
Ко времени менархе в яичнике содержится около 200
-400 тыс. фолликулов, из которых созревают до
стадии овуляции 300-400.ПРИМОРДИАЛЬНЫЙ ФОЛЛИКУЛ (а)
Состоит из незрелой яйцеклетки, которая
расположена в фолликулярном и гранулезном
(зернистом) эпителии.
Снаружи фолликул окружен соединительной
оболочкой (текаклетки).В течение
1 - овоцит, 2 - гранулезные клетки (зернистая зона)
каждого менструаль
ного цикла от 3 до
30 примордиальных
фолликулов начина
ют рости, и из них
формируются преан
тральные, или пер
вичные, фолликулы.ПРЕАНТРАЛЬНЫЙ ФОЛЛИКУЛ (б)
Овоцит увеличивается и окружается мембраной,
называемой блестящей оболочкой (zona
pellucida). Клетки гранулезного эпителия
подвергаются размножению, а слой теки
образуется из окружающей стромы.
Клетки гранулезного
слоя преантрального
фолликула способны
синтезировать стерои
ды трех классов, при
этом эстрогенов син
тезируется гораздо
больше, чем андроге
нов и прогестерона.АНТРАЛЬНЫЙ, или ВТОРИЧНЫЙ ФОЛЛИКУЛ (в)
Характеризуется дальнейшим ростом:
увеличивается число клеток гранулезного слоя,
продуцирующих фолликулярную жидкость.
Фолликулярная жидкость накапливается в
межклеточном пространстве гранулезного слоя
и образует полости.
1 - овоцит, 2 - гранулезные клетки (зернистая зона),
В этот период фоллику 3 - тека-клетки, 4 - базальная мембрана
логенеза (8-9-й день
менструального
цикла) отмечается син
тез половых стероидных
гормонов: эстрогенов и
андрогенов.АНТРАЛЬНЫЙ, или ВТОРИЧНЫЙ ФОЛЛИКУЛ
Согласно современной теории синтеза половых
гормонов
В тека-клетках синтезируются андрогены -
андростендион и тестостерон.
Затем андрогены попадают в клетки
гранулезного слоя, и в них ароматизируются в
эстрогены.ДОМИНАНТНЫЙ ФОЛЛИКУЛ (г)
Как правило, один такой фолликул образуется из
множества антральных фолликулов (к 8-му дню цикла).
Он является самым крупным, содержит наибольшее
число клеток гранулезного слоя и рецепторов к ФСГ, ЛГ.
Доминантный фолликул
имеет богато васкуляризи
рованный тека-слой.
Наряду с ростом и развити
ем доминантного преовуля
торного фолликула в яич
никах параллельно проис
ходит процесс атрезии
остальных (90 %) растущих
фолликулов.
1 - овоцит, 2 - гранулезные клетки (зернистая зона),
3 - тека-клетки, 4 - базальная мембранаДОМИНАНТНЫЙ ФОЛЛИКУЛ
Доминантный фолликул в первые дни
менструального цикла имеет диаметр 2 мм,
который в течение 14 дней к моменту овуляции
увеличивается в среднем до 21 мм.
За это время происходит 100-кратное
увеличение объема фолликулярной жидкости.
В ней резко возрастает содержание эстрадиола
и ФСГ, а также определяются факторы роста.ОВУЛЯЦИЯ
Разрыв преовулярного доминантного (третичного)
фолликула и выход из него яйцеклетки.
Ко времени овуляции в овоците происходит процесс
мейоза.
Овуляция сопровождается кровотечением из
разрушенных капилляров, окружающих тека-клетки.
Полагают, что овуляция происходит через 24-36 ч
после формирования преовуляторного пика
эстрадиола.
Истончение и разрыв стенки преовуляторного
фолликула происходят под влиянием фермента
коллагеназы, простагландинов F2 и Е2,
протеолитических ферментов, образующихся в
гранулезных клетках, окситоцина и релаксина.ЖЕЛТОЕ ТЕЛО (corpus luteum)
После выхода яйцеклетки фолликул
претерпевает специфические изменения,
заканчивающиеся образованием желтого тела.
Желтое тело - транзиторная эндокринная
железа, которая функционирует в течение 14
дней независимо от продолжительности
менструального цикла.
При отсутствии бере
менности желтое тело
регрессирует.ЖЕЛТОЕ ТЕЛО (corpus luteum)
1) стадия пролиферации и лютеинизации
гранулезных клеток (в клетках гранулезы
накапливается липохромный пигмент лютеин, и
они преобразуются в лютеиновые и текалютеиновые клетки);
2) стадия васкуляризации (появление богатой
кровеносной сети, сосуды которой направляются
от внутренней зоны к центру желтого тела);
Стадии формирования и развития ЖТ:
3) стадия расцвета (период максимального
развития и функционирования - на 21–22-й день
менструального цикла или через 6–8 дней после
пика ЛГ; определяет второй пик эстрогенов и пик
прогестерона);
4) стадию угасания - в лютеиновых клетках
доминируют дистрофические процессы, желтое
тело фиброзируется и гиалинизируется, его
размеры уменьшаются; впоследствии, через 1–2
мес, на месте желтого тела формируется белое
тело (corpus albicans), которое затем полностью
рассасывается.АНАТОМИЯ ЯИЧНИКААНАТОМИЯ ЯИЧНИКА
Примордиальный
фолликул
Первичный фолликул
Антральный фолликул
Атретический фолликул
Зрелый фолликул
Овуляция
Желтое тело
(ранняя стадия)
Желтое тело
(расцвет)
Зародышевый
эпителий
Белое тело
БрыжейкаЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ
Таким образом, в яичнике синтезируются основные
женские половые стероидные гормоны - эстрадиол и
прогестерон, а также андрогены.
В I фазу менструального цикла, которая длится от первого
дня менструации до момента овуляции, организм находится
под влиянием эстрогенов.
Во II (от овуляции до начала менструации) к эстрогенам
присоединяется прогестерон, выделяющийся клетками
желтого тела.
Первая фаза менструального цикла называется также
фолликулиновой, или фолликулярной, вторая фаза цикла -
лютеиновой.ОРГАНЫ- МИШЕНИ
Матка, шейка матки, влагалище, молочные
железы.
Губчатое вещество костей, мозг, эндотелий и
гладкие мышечные клетки сосудов, миокард,
уретра, оболочки глаза.
Кожа и ее придатки (волосяные фолликулы и
сальные железы) и др.ОРГАНЫ- МИШЕНИ
Биологический эффект дают свободные
(несвязанные) фракции эстрадиола и
тестостерона (1 %).
Основная масса яичниковых гормонов (99%)
находится в связанном состоянии.
Транспорт осуществляется специальными
белками - стероидосвязывающими глобулинами
и неспецифическими транспортными системами
- альбуминами и эритроцитами.
Секретируются клетками зернистой
мембраны, внутренней оболочки и в меньшей
степени интерстициальными клетками.
В незначительном количестве эстрогены
образуются в желтом теле, корковом слое
надпочечников, у беременных - в плаценте
(синцитиальных клетках хориальных ворсин)ЭСТРОГЕНЫ (Эстрадиол, эстриол, эстрон)
Стимулируют развитие вторичных половых
признаков
Вызывают гиперплазию и гипертрофию
эндометрия и миометрия
Улучшают кровоснабжение матки
Способствуют развитию выводящей системы
молочных желез
В физиологическом количестве стимулируют
ретикулоэндотелиальную систему (усиливают
выработку антител и активность фагоцитов, повышая
устойчивость организма к инфекциям)
Задерживают в мягких тканях азот, натрий, жидкость,
в костях - кальций, фосфор
Вызывают увеличение концентрации гликогена,
глюкозы, фосфора, креатинина, железа и меди в
крови и мышцах
Снижают содержание холестерина, фосфолипидов и
общего жира в печени и крови, ускоряют синтез
высших жирных кислот.
Секретируются лютеиновыми клетками желтого
тела
Лютеинизирующими клетками гранулезы и
оболочек фолликула (основной источник вне
беременности)
А также корковым веществом надпочечников и
плацентой.ГЕСТАГЕНЫ (прогестерон, 17α-оксипрогестерон, D4прегненол-20α-он-3, D4-прегненол-20β-он-3)
Подавляют
пролиферацию эндометрия, вызванную
эстрогенами
Осуществляют секреторные преобразования в
эндометрии
При оплодотворении яйцеклетки гестагены
ингибируют овуляцию
Препятствуют сокращению матки («протектор»
беременности)
Способствуют развитию альвеол в молочных
железах.ГЕСТАГЕНЫ (прогестерон, 17α-оксипрогестерон,
D4-прегненол-20α-он-3, D4-прегненол-20β-он-3)
Вызывают возбуждение терморегулирующего
центра, локализованного в гипоталамусе, что
проявляется повышением базальной температуры
В физиологических условиях уменьшают
содержание аминного азота в плазме крови,
увеличивают экскрецию аминокислот, усиливают
отделение желудочного сока, тормозят отделение
желчи
дегидроэпиандростерон, тестостерон и
эпитестостерон)
Андрогены секретируются клетками внутренней
оболочки фолликула, интерстициальными клетками (в
незначительном количестве) и в сетчатой зоне
коркового вещества надпочечников (основной
источник).
– Вирильный эффект - большие дозы андрогенов
вызывают гипертрофию клитора, оволосение по
мужскому типу, разрастание перстневидного хряща,
появление acne vulgaris.
– Гонадотропный эффект - малые дозы андрогенов
стимулируют секрецию гонадотропных гормонов,
способствуют росту и созреванию фолликула,
овуляции, лютеинизации.
- Антигонадотропный эффект - высокий уровень
концентрации андрогенов в предовуляторном
периоде подавляет овуляцию и в дальнейшем
вызывает атрезию фолликула.
– Эстрогенный эффект - в малых дозах
андрогены вызывают пролиферацию эндометрия
и эпителия влагалища.
- Антиэстрогенный эффект - большие дозы
андрогенов блокируют процессы пролиферации в
эндометрии и приводят к исчезновению
ацидофильных клеток во влагалищном мазке.АНДРОГЕНЫ (Андростендион, дегидроэпиандростерон,
тестостерон и эпитестестерон)
Обладают выраженной анаболической
активностью, усиливают синтез белка
тканями;
задерживают в организме азот, натрий и
хлор, снижают выведение мочевины.
Ускоряют рост костей и окостенение
эпифизарных хрящей;
увеличивают количество эритроцитов и
гемоглобина.Другие гормоны яичников:
Ингибин, синтезируемый зернистыми
клетками, оказывает тормозящее воздействие
на синтез ФСГ;
окситоцин (обнаружен в фолликулярной
жидкости, желтом теле) - в яичниках
обладает лютеолитическим воздействием,
способствует регрессу желтого тела;
релаксин - образуется в клетках гранулезы и
желтом теле, способствует овуляции,
расслабляет миометрий.МАТОЧНЫЙ ЦИКЛ
Под влиянием гормонов яичников в миометрии
и эндометрии наблюдаются циклические
изменения, соответствующие фолликулиновой
и лютеиновой фазам в яичниках.
Мышечный слой
Для фолликулиновой
фазы –
гипертрофия клеток
Для лютеиновой фазы –
гиперплазия
клетокМАТОЧНЫЙ ЦИКЛ
Эндометрий
Состоит из следующих слоев
1.Базальный слой, который не оттор
гается во время менструации. Из его
клеток в течение менструального цикла об
разуется слой эндометрия.
2. Поверхностный слой, состоящий из
компактных эпителиальных клеток,
которые выстилают полость матки.
3. Промежуточный, или спонгиозный,
слой.
Последние два слоя составляют
функциональный слой,
подвергающийся основным циклическим изменениям в течение
менструального цикла и отторгающийся в период менструации.МАТОЧНЫЙ ЦИКЛ
Фазы изменения
эндометрия в течение
цикла:
1)фаза пролиферации
(ранняя, средняя, поздняя);
2) фаза секреции
(ранняя, средняя, поздняя);
3) фаза десквамации
(менструация)
4) фаза регенерацииФАЗА ПРОЛИФЕРАЦИИ
По мере увеличения секреции
эстрадиола растущими фолли
кулами яичников
эндометрий претерпевает
пролиферативные изменения.
Происходит активное
размножение клеток
базального слоя.
Образуется новый
поверхностный рыхлый слой с
вытянутыми трубчатыми
железами.
Этот слой быстро утолщается
в 4-5 раз. Трубчатые железы,
выстланные цилиндрическим
эпителием, удлиняются.
Различают раннюю, среднюю и
позднюю фазы пролиферации.ФАЗА СЕКРЕЦИИ
В лютеиновую фазу яичниково
Го цикла под влиянием прогес
терона увеличивается извилис
тость желез, а просвет их пос
тепенно расширяется.
Клетки стромы, увеличиваясь
в объеме, приближаются друг
к другу.
Секреция желез усиливается.
В просвете желез находят
обильное количество секрета.
В зависимости от интенсивнос
ти секреции железы либо оста
ются сильно извитыми, либо
приобретают пилообразную
форму.
Отмечается усиленная васкуля
ризация стромы.
Различают раннюю, среднюю и позднюю фазы секреции.
Это отторжение функционального слоя эндометрия.
Установлено, что эндокринной основой начала
менструации является выраженное снижение уровней
прогестерона и эстрадиола вследствие регрессии
желтого тела.МЕНСТРУАЦИЯ ИЛИ ФАЗА КРОВОТЕЧЕНИЯ И ДЕСКВАМАЦИИ
Существуют следующие основные локальные механизмы,
принимающие участие в менструации:
1) спазм спиральных артериол (F2α), ломкость и
проницаемость сосудистой стенки;
2) изменение механизмов гемостаза в матке (стаз,
образование тромбов);
3) ишемия тканей, кровоизлияние в строму, лейкоцитарная
инфильтрация
4) усиленное выделение ферментов лизосом клетками
эндометрия;
5) некробиоз ткани и ее расплавление;
6) расширение сосудов (F2) после длительного спазма,
полнокровие, разрыв сосудов и отторжение
некротизированных отделов эндометрия.ФАЗА РЕГЕНЕРАЦИИ
Регенерация эндометрия
Наблюдается с самого начала менструации.
К концу 24-го часа менструации отторгается 2/з
функционального слоя эндометрия.
Базальный слой содержит эпителиальные
клетки стромы, являющиеся основой для
регенерации эндометрия, которая обычно к 5-му
дню цикла полностью завершается.
Параллельно завершается ангиогенез с
восстановлением целости разорванных
артериол, вен и капилляров.РЕГУЛЯЦИЯ МЕНСТРУАЛЬНОГО ЦИКЛАМЕНСТРУАЛЬНЫЙ ЦИКЛ
СИСТЕМЫ
В основе лежит цирхоральная секреция и
выделение ГТ-РГ гипоталамусом, а также
регуляция синтеза и выделения
гонадотропинов эстрадиолом и ингибином по
мехнизмам отрицательной и положительной
обратной связи.
Взаимосвязь всех звеньев репродуктивнои
системы обеспечивается наличием в них
рецепторов как к половым, так и к
гонадотропным гормонам.ФУНКЦИОНИРОВАНИЕ РЕПРОДУКТИВНОЙ
СИСТЕМЫ
Выделяют следующие типы обратной связи:
1)"длинная петля" обратной связи - между
гормонами яичника и ядрами гипоталамуса;
между гормонами яичника и гипофизом;
2)"короткая петля" - между передней долей
гипофиза и гипоталамусом;
3) "ультракороткая петля" - между ГТРГ и
нервными клетками гипоталамуса.ФУНКЦИОНИРОВАНИЕ РЕПРОДУКТИВНОЙ
СИСТЕМЫ
У женщины репродуктивного возраста имеется
как отрицательная, так и положительная
обратная связь между яичниками и гипоталамогипофизарной системой.
Примером отрицательной обратной связи
является усиление выделения ЛГ и ФСГ передней
долей гипофиза в ответ на низкий уровень
эстрадиола в раннюю фолликулярную фазу цикла.
Примером положительной обратной связи
является выброс ЛГ и ФСГ в ответ на овуляторный
максимум содержания эстрадиола в крови.ФУНКЦИОНИРОВАНИЕ РЕПРОДУКТИВНОЙ
СИСТЕМЫ
Другим примером отрицательной обратной
связи может быть усиление образования ГТ-РГ
при снижении концентрации ЛГ в клетках
передней доли гипофиза, или, напротив,
подавление секреции гормонов гипофиза
(гипоталамуса) в ответ на повышение уровня
секреции соответствующих гормонов яичников
(гипофиза).
РЕПРОДУКТИВНОЙ СИСТЕМЫ
РГЛГ - рилизинггормоны;
ОК - окситоцин,
Прл - пролактин;
ФСГ -
фолликулостимулирую
щий гормон;
П - прогестерон; Э -
эстрогены;
А - андрогеиы; Р -
релаксин;
И - ингибин;
ЛГ - лютеинизирующий
гормон.КРИТЕРИИ НОРМАЛЬНОГО МЕНСТРУАЛЬНОГО
ЦИКЛА:
двухфазность;
продолжительность не менее 21 и не более 35
дней
(28±7 дней; у 60% женщин - 28 дней);
цикличность;
продолжительность менструации - 2–7 дней
(4±2 дня);
менструальная кровопотеря - до 80 мл;
отсутствие болезненности и нарушений общего
состояния.ТЕСТЫ ФУНКЦИОНАЛЬНОЙ ДИАГНОСТИКИ
базальная температура;
симптом «зрачка»;
кариопикнотический индекс;
симптом натяжения слизи;
симптом папоротника.БАЗАЛЬНАЯ ТЕМПЕРАТУРА
Измеряется в прямой кишке утром, до подъема с постели.
При овуляторном менструальном цикле базальная
температура повышается в лютеиновую фазу цикла на 0,4-
0,6 °С и держится в течение всей второй фазы.
В день менструации или за день до нее базальная
температура снижается.
При беременности повышение базальной температуры
объясняется возбуждением терморегулирующего центра
гипоталамуса под влиянием прогестерона.СИМПТОМ ЗРАЧКА
Симптом зрачка отражает изменения в слизи
шейки матки. Под влиянием эстрогенов в шейке
матки накапливается прозрачная стекловидная
слизь, что обусловливает расширение наружного
отверстия шейки матки.
Максимальное количество слизи наблюдается в
предовуляторные дни цикла, наружное отверстие
становится темным, напоминает зрачок.
Во вторую фазу цикла под влиянием прогестерона
количество слизи уменьшается или же она
полностью исчезает. Слизь имеет глыбчатое
строение.
Различают 3 степени симптома зрачка:
+, + + , + + + .КАРИОПИКНОТИЧЕСКИЙ ИНДЕКС
Кариопикнотический индекс. Под влиянием гормонов
яичников также происходят циклические изменения в
слизистой оболочке влагалища, особенно в его верхней
трети.
В мазке из влагалища могут встречаться следующие виды
клеток плоского многослойного эпителия: а) ороговевающие,
б) промежуточные, в) базальные, или атрофические.
Клетки первого типа начинают преобладать по мере
нарастания секреции яичниками эстрогенов.
На основании определения количественных соотношений
клеточных элементов можно судить о степени насыщенности
организма эстрогенными гормонами или об их
недостаточности.
Максимальное число ороговевающих клеток выявляется в
предовуляторные дни - 80-88 %,
в раннюю фазу пролиферации - 20-40 %,
в позднюю фазу секреции - 20-25 %.СИМПТОМ НАТЯЖЕНИЯ СЛИЗИ
Симптом натяжения слизи. Корнцангом
берут слизь шеечного канала и путем
разведения браншей определяют ее
эластичность (растяжимость).
Натяжение слизи более 6-8 см
свидетельствует о достаточной эстрогенной
насыщенности.СИМПТОМ «ЛИСТА ПАПОРОТНИКА»
Симптом «листа папоротника» основан на
кристаллизации шеечной слизи, нанесенной на
предметное стекло.
После обнажения с помощью зеркал шейки матки
пинцетом или корнцангом берут содержимое
шеечного канала и наносят его на предметное стекло
в виде кружка. Кристаллизация солей шеечной слизи
происходит в присутствии муцина под действием
эстрогенов.
Симптом «листа папоротника»можно установить
между 7-м и 20-21 днем нормального
менструального цикла; наивысшего развития
симптом достигает к моменту овуляции.
Степень выраженности симптома обозначают
знаками:
+, + + , + + + .Циклические изменения в
органах репродуктивной
системы в течение
менструального цикла
I - гонадотропная
регуляция функции
яичников; ПДГ - передняя
доля гипофиза; II -
содержание в эндометрии
реиепторов к эстрадиолу
- РЭ (1,2,3; сплошная
линия) и прогестерону - РП
(2,4,6; пунктирная линия);
II - циклические изменения
эндометрия; IV -
цитология эпителия
влагалища; V - базальная
температура; VI -
натяжение цервикапьной
слизи.
11020 0
Менструация (от menstruus — месячный) - циклическое непродолжительное маточное кровотечение - отражает несостоятельность сложной интегрированной системы, предназначенной для обеспечения зачатия и развития беременности на ранних стадиях. Эта система включает высшие мозговые центры, гипоталамус, гипофиз, яичники, матку и органы мишени, функционально связанные между собой. Комплекс сложных биологических процессов, происходящих в период между менструациями, именуется менструальным циклом, длительность которого принято отсчитывать от первого дня предыдущего до первого дня последующего кровотечения. Продолжительность менструального цикла в норме колеблется от 21 до 36 дней, наиболее часто наблюдается 28-дневный менструальный цикл; длительность менструального кровотечения варьирует от 3 до 7 дней, объем кровопотери не превышает 100 мл.
Кора головного мозга
Регуляция нормального менструального цикла происходит на уровне специализированных нейронов головного мозга, которые получают информацию о состоянии внешней среды, преобразуют ее в нейрогормональные сигналы. Последние, в свою очередь, через систему нейротрансмиттеров (передатчиков нервных импульсов) поступают в нейросекреторные клетки гипоталамуса. Функцию нейротрансмиттеров выполняют биогенные амины-катехоламины - дофамин и норадреналин, индолы - серотонин, а также нейропептиды морфиноподобного происхождения, опиоидные пептиды - эндорфины и энкефалины.
Дофамин, норадреналин и серотонин осуществляют контроль над гипоталамическими нейронами, секретирующими гонадотропин-рилизинг-фактор (ГТРФ): дофамин поддерживает секрецию ГТРФ в аркуатных ядрах, а также тормозит выделение аденогипофизом пролактина; норадреналин регулирует передачу импульсов в преоптические ядра гипоталамуса и стимулирует овуляторный выброс ГТРФ; серотонин контролирует циклическую секрецию ГТРФ из нейронов переднего (зрительного) отдела гипоталамуса. Опиоидные пептиды подавляют секрецию лютеинизирующего гормона, угнетают стимулирующее действие дофамина, а их антагонист, налаксон, вызывает резкий подъем уровня ГТРФ.
Гипоталамус
Ядра гипофизотропной зоны гипоталамуса (супраоптические, паравентрикулярные, аркуатные и вентромедиальные) вырабатывают специфические нейросекреты с диаметрально противоположным фармакологическим эффектом: либерины, или рилизинг-факторы (realizing factors), освобождающие соответствующие тройные гормоны в передней доле гипофиза и статины, ингибирующие их выделение.
В настоящее время известны семь либеринов - кортиколиберин (адренокортикотропный рилизинг-фактор, АКТГ-РФ), соматолиберин (соматотропный СТГ-РФ), тиреолиберин (тиреотропный рилизинг-фактор, Т-РФ), меланолиберин (меланотропный рилизинг-фактор, М-РФ), фоллиберин (фолликулостимулирующий рилизинг-фактор, ФСГ-РФ), люлиберин (лютеинизирующий рилизинг-фактор, ЛГ-РФ), пролактолиберин (пролактин рилизинг-фактор, ПРФ) и три статина - меланостатин (меланотропный ингибирующий фактор, М-ИФ), соматостатин (соматотропный ингибирующий фактор, С-ИФ), пролактостатин (пролактин ингибирующий фактор, ПИФ).
Лютеионизирующий рилизинг-фактор выделен, синтезирован и подробно описан. Вместе с тем, химическая природа фоллиберина и его аналогов до настоящего времени не изучена. Однако доказано, что люлиберин обладает способностью стимулировать секрецию обоих гормонов аденогипофиза - как фолликулостимулирующего, так и лютеинизирующего гормонов. Поэтому для этих либеринов общепризнанным является термин - гонадолиберин, или гонадотропин-рилизинг-фактор (ГТРФ).
Кроме гипофизотропных гормонов, супраоптические и паравентрикулярные ядра гипоталамуса синтезируют два гормона - вазопрессин (антидиуретический гормон, АДГ) и окситоцин, которые депонируются в нейрогипофизе.
Гипофиз
Базофильные клетки аденогипофиза - гонадотропоциты - выделяют гормоны - гонадотропины, принимающие непосредственное участие в регуляции менструального цикла. К гонадотропным гормонам относятся фоллитропин, или фолликулостимулирующий гормон (ФСГ) и лютропин, или лютеинизирующий гормон (ФСГ). Лютропин и фоллитропин представляют гликопротеины, состоящие из двух пептидных цепей - а- и b-субъединиц; a-цепи гонадотропинов идентичны, в то время как различие b-звеньев определяет их биологическую специфичность.
ФСГ стимулирует рост и созревание фолликулов, пролиферацию клеток гранулезы, а также индуцирует образование рецепторов ЛГ на поверхности этих клеток. Под влиянием ФСГ повышается уровень ароматаз в зреющем фолликуле. Лютропин оказывает воздействие на синтез андрогенов (предшественников эстрогенов) в тека-клетках, в комплексе с ФСГ обеспечивает овуляцию и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула. В настоящее время обнаружено два типа секреции гонадотропинов - тонический и циклический. Тоническое выделение гонадотропинов способствует развитию фолликулов и продукции ими эстрогенов; циклическое - обеспечивает смену фаз низкой и высокой секреции гормонов и, в частности, их предовуляторный пик.
Группа ацидофильных клеток передней доли гипофиза - лактотропоциты - продуцирует пролактин (ПРЛ). Пролактин образован одной пептидной цепью, биологическое действие его многообразно:
1) ПРЛ стимулирует рост молочных желез и регулирует лактацию;
2) обладает жиромобилизующим и гипотензивным эффектом;
3) в повышенных количествах обладает ингибирующим воздействием на рост и созревание фолликула.
Другие гормоны аденогипофиза (тиротропин, кортикотропин, соматотропин, меланотропин) играют второстепенную роль в генеративных процессах человека.
Задняя доля гипофиза, нейрогипофиз, как указывалось выше, не является эндокринной железой, а лишь депонирует гормоны гипоталамуса - вазопрессин и окситоцин, которые находятся в организме в виде белкового комплекса (белок Ван Дейка).
Яичники
Генеративная функция яичников характеризуется циклическим созреванием фолликула, овуляцией, выделением яйцеклетки, способной к зачатию, и обеспечением секреторных преобразований в эндометрии, направленных для восприятия оплодотворенной яйцеклетки.
Основной морфофункциональной единицей яичников является фолликул. В соответствии с Международной гистологической классификацией (1994) выделяют 4 типа фолликулов: примордиальные, первичные, вторичные (антральные, полостные, пузырчатые), зрелые (преовуляторные, граафовы).
Примордиальные фолликулы образуются на пятом месяце внутриутробного развития плода и существуют в течение нескольких лет после стойкого прекращения менструаций. К моменту рождения в обоих яичниках содержится около 300 000-500 000 примордиальных фолликулов, в дальнейшем их число резко снижается и к 40 годам составляет около 40 000-50 000 (физиологическая атрезия примордиальных фолликулов). Примордиальный фолликул состоит из яйцеклетки, окруженной одним рядом фолликулярного эпителия; диаметр его не превышает 50 мкм (рис. 1).
Рис. 1. Анатомия яичника
Стадия первичного фолликула характеризуется усиленным размножением фолликулярного эпителия, клетки которого приобретают зернистое строение и образуют зернистый (гранулезный) слой (stratum granulosum). Клетки этого слоя выделяют секрет (liquor folliculi), который скапливается в межклеточном пространстве. Величина яйцеклетки постепенно увеличивается до 55-90 мкм в диаметре. Образовавшаяся жидкость оттесняет яйцеклетку к периферии, где клетки зернистого слоя окружают ее со всех сторон и образуют яйценосный бугорок (cumulus oophorus). Другая часть этих клеток смещается к периферии фолликула и формирует тонкослойную зернистую (гранулезную) мембрану (membrana granulosus).
В процессе формирования вторичного фолликула происходит растяжение жидкостью его стенок: овоцит в этом фолликуле уже не увеличивается (к данному моменту диаметр его составляет 100-180 мкм), однако диаметр самого фолликула возрастает и достигает 10-20 мм. Оболочка вторичного фолликула отчетливо дифференцирована на наружную и внутреннюю. Внутренняя оболочка (theca interna), состоит из 2-4 пластов клеток, расположенных на зернистой мембране. Наружная оболочка (theca externa), локализуется непосредственно на внутренней и представлена дифференцированной соединительнотканной стромой.
В зрелом фолликуле яйцеклетка, заключенная в яйценосный бугорок, покрыта прозрачной (стекловидной) оболочкой (zona pellucida), на которой зернистые клетки располагаются в радиальном направлении и образуют лучистый венец (corona radiata) (рис. 2).

Рис. 5. Развитие фолликула
Овуляция - разрыв зрелого фолликула с выходом яйцеклетки, окруженной лучистым венцом, в брюшную полость, а в дальнейшем и в ампулу маточной трубы. Нарушение целостности фолликула происходит в наиболее тонкой и выпуклой его части, именуемой стигмой (stigma folliculi).
Созревание фолликула происходит периодически, через определенный интервал времени. У приматов и человека в течение менструального цикла созревает один фолликул, остальные подвергаются обратному развитию и превращаются в фиброзные и атретические тельца. В течение всего репродуктивного периода овулирует около 400 яйцеклеток, остальные овоциты подвергаются атрезии. Жизнеспособность яйцеклетки находится в пределах 12-24 часов.
Лютеинизация представляет специфические преобразования фолликула в постовуляторном периоде. В результате лютеинизации (окрашивания в желтый цвет вследствие накопления липохромного пигмента - лютеина), размножения и разрастания клеток зернистой мембраны овулировавшего фолликула формируется образование, именуемое желтым телом (corpus luteum) (лютеинизации подвергаются также клетки внутренней зоны, трансформирующиеся в тека-клетки). В тех случаях, когда оплодотворения не происходит, желтое тело существует 12-14 дней и претерпевает следующие стадии развития:
а) стадия пролиферации характеризуется разрастанием гранулезных клеток и гиперемией внутренней зоны;
б) стадия васкуляризации отличается появлением богатой кровеносной сети, сосуды которой направляются от внутренней зоны к центру желтого тела; размножающиеся гранулезные клетки превращаются в полигональные, в протоплазме которых накапливается лютеин;
в) стадия расцвета - период максимального развития, лютеиновый слой приобретает специфичную для желтого тела складчатость;
г) стадия обратного развития - наблюдается дегенеративная трансформация лютеиновых клеток, желтое тело обесцвечивается, фиброзируется и гиалинизируется, размеры его непрерывно уменьшаются; впоследствии, через 1-2 месяца, на месте желтого тела формируется белое тело (corpus albicans), которое затем полностью рассасывается.
Таким образом, яичниковый цикл состоит из двух фаз - фолликулиновой и лютеиновой. Фолликулиновая фаза начинается после менструации и оканчивается овуляцией; лютеиновая фаза занимает промежуток между овуляцией и началом менструации.
Гормональная функция яичников
Клетки гранулезной мембраны, внутренней оболочки фолликула и желтого тела в период своего существования выполняют функцию железы внутренней секреции и синтезируют три основных типа стероидных гормонов - эстрогены, гестагены, андрогены.
Эстрогены секретируются клетками зернистой мембраны, внутренней оболочки и в меньшей степени интерстициальными клетками. В незначительном количестве эстрогены образуются в желтом теле, корковом слое надпочечников, у беременных - в плаценте (синцитиальных клетках хориальных ворсин). Основными эстрогенами яичника являются эстрадиол, эстрон и эстриол (преимущественно синтезируются первые два гормона).
Активность 0,1 мг эстрона условно принята за 1 ME эстрогенной активности. По тесту Аллена и Дойзи (наименьшее количество препарата, вызывающее течку у кастрированных мышей) наибольшей активностью обладает эстрадиол, далее эстрон и эстриол (соотношение 1:7:100).
Метаболизм эстрогенов
Эстрогены циркулируют в крови в свободном и связанном с белком (биологические неактивные) виде. Основное количество эстрогенов находится в плазме крови (до 70%), 30% - в форменных элементах. Из крови эстрогены поступают в печень, затем в желчь и кишечник, откуда частично снова всасываются в кровь и проникают в печень (энтерогепатический кругооборот), частично - выводятся с калом. В печени эстрогены инактивируются путем образования парных соединений с серной и глюкуроновой кислотами, которые поступают в почки и выводятся с мочой.
Воздействие стероидных гормонов на организм систематизируется следующим образом.
Вегетативное воздействие (строго специфично) - эстрогены оказывают специфическое действие на женские половые органы: стимулируют развитие вторичных половых признаков, вызывают гиперплазию и гипертрофию эндометрия и миометрия, улучшают кровоснабжение матки, способствуют развитию выводящей системы молочных желез.
Генеративное воздействие (менее специфично) - эстрогены стимулируют трофические процессы в период созревания фолликула, способствуют формированию и росту гранулезы, образованию яйцеклетки и развитию желтого тела; подготавливают яичник к воздействию гонадотропных гормонов.
Общее воздействие (неспецифично) - эстрогены в физиологическом количестве стимулируют ретикулоэндотелиальную систему (усиливают выработку антител и активность фагоцитов, повышая устойчивость организма к инфекциям), задерживают в мягких тканях азот, натрий, жидкость, в костях - кальций, фосфор. Вызывают увеличение концентрации гликогена, глюкозы, фосфора, креатинина, железа и меди в крови и мышцах; снижают содержание холестерина, фосфолипидов и общего жира в печени и крови, ускоряют синтез высших жирных кислот.
Гестагены секретируются лютеиновыми клетками желтого тела, лютеинизирующими клетками гранулезы и оболочек фолликула (основной источник вне беременности), а также корковым веществом надпочечников и плацентой. Основной гестаген яичников - прогестерон, помимо прогестерона яичники синтезируют 17а-оксипрогестерон, D4-прегненол-20а-он-3, D4-прегненол-20b-он-3.
Метаболизм гестагенов протекает по схеме: прогестерон-аллопрегнанолон-прегнанолон-прегнандиол. Последние два метаболита не обладают биологической активностью: связываясь в печени с глюкуроновой и серной кислотами, выделяются с мочой.
Вегетативное воздействие - гестагены оказывают влияние на половые органы после предварительной эстрогенной стимуляции: подавляют пролиферацию эндометрия, вызванную эстрогенами, осуществляют секреторные преобразования в эндометрии; при оплодотворении яйцеклетки гестагены подавляют овуляцию, препятствуют сокращению матки («протектор» беременности), способствуют развитию альвеол в молочных железах.
Генеративное воздействие - гестагены в малых дозах стимулируют секрецию ФСГ, в больших - блокируют как ФСГ, гак и ЛГ; вызывают возбуждение терморегулирующего центра, локализованного в гипоталамусе, что проявляется повышением базальной температуры.
Общее воздействие - гестагены в физиологических условиях уменьшают содержание аминного азота в плазме крови, увеличивают экскрецию аминокислот, усиливают отделение желудочного сока, тормозят отделение желчи.
Андрогены секретируются клетками внутренней оболочки фолликула, интерстициальными клетками (в незначительном количестве) и в сетчатой зоне коркового вещества надпочечников (основной источник). Основные андрогены яичников - андростендион и дсгидроэпиандростерон, в малых дозах синтезируются тестостерон и эпитестестерон.
Специфическое действие андрогенов на репродуктивную систему зависит от уровня их секреции (малые дозы стимулируют функцию гипофиза, большие - ее блокируют) и может проявляться в виде следующих эффектов:
- вирильный эффект - большие дозы андрогенов вызывают гипертрофию клитора, оволосение по мужскому типу, разрастание перстневидного хряща, появление acne vulgaris;
- гонадотропный эффект - малые дозы андрогенов стимулируют секрецию гонадотропных гормонов, способствуют росту и созреванию фолликула, овуляции, лютеинизации;
- антигонадотропный эффект - высокий уровень концентрации андрогенов в предовуляторном периоде подавляет овуляцию и в дальнейшем вызывает атрезию фолликула;
- эстрогенный эффект - в малых дозах андрогены вызывают пролиферацию эндометрия и эпителия влагалища;
- антиэстрогенный эффект - большие дозы андрогенов блокируют процессы пролиферации в эндометрии и приводят к исчезновению ацидофильных клеток во влагалищном мазке.
Общее воздействие
Андрогены обладают выраженной анаболической активностью, усиливают синтез белка тканями; задерживают в организме азот, натрий и хлор, снижают выведение мочевины. Ускоряют рост костей и окостенение эпифизарных хрящей, увеличивают количество эритроцитов и гемоглобина.
Другие гормоны яичников: ингибин, синтезируемый зернистыми клетками, оказывает тормозящее воздействие на синтез ФСГ; окситоцин (обнаружен в фолликулярной жидкости, желтом теле) - в яичниках обладает лютеолитическим воздействием, способствует регрессу желтого тела; релаксин, образуется в клетках гранулезы и желтом теле, способствует овуляции, расслабляет миометрий.
Матка
Под влиянием гормонов яичников в миометрии и эндометрии наблюдаются циклические изменения, соответствующие фолликулиновой и лютеиновой фазам в яичниках. Для фолликулиновой фазы характерна гипертрофия клеток мышечного слоя матки, для лютеиновая - их гиперплазия. Функциональные изменения в эндометрии отражаются последовательной сменой стадий пролиферации, секреции, десквамации (менструации), регенерации.
Фаза пролиферации (соответствует фолликулиновой фазе) характеризуется преобразованиями, возникающими под влиянием эстрогенов.
Ранняя стадия пролиферации (до 7-8 дня менструального цикла): поверхность слизистой оболочки выстлана уплощенным цилиндрическим эпителием, железы имеют вид прямых или слегка извитых коротких трубок с узким просветом, эпителий желез однорядный низкий цилиндрический; строма состоит из веретенообразных или звездчатых ретикулярных клеток с нежными отростками, в клетках стромы и эпителия - единичные митозы.
Средняя стадия пролиферации (до 10-12 дня менструального цикла): поверхность слизистой оболочки выстлана высоким призматическим эпителием, железы удлиняются, становятся более извитыми, строма отечна, разрыхлена; увеличивается количество митозов.
Поздняя стадия пролиферации (до овуляции): железы становятся резко извитыми, иногда шпорообразными, просвет их расширяется, эпителий, выстилающий железы, - многорядный, строма сочная, спиральные артерии достигают поверхности эндометрия, умеренно извиты.
Фаза секреции (соответствует лютеиновой фазе) отражает изменения, обусловленные воздействием прогестерона.
Ранняя стадия секреции (до 18 дня менструального цикла) характеризуется дальнейшим развитием желез и расширением их просвета, наиболее характерный признак этой стадии - появление в эпителии субнуклеарных вакуолей, содержащих гликоген; митозы в эпителии желез в конце стадии отсутствуют; строма сочная, рыхлая.
Средняя стадия секреции (19-23 день менструального цикла) - отражает преобразования, характерные для периода расцвета желтого тела, т. е. периода максимальной гестагенной насыщенности. Функциональный слой становится более высоким, отчетливо разделяется на глубокий и поверхностный слои: глубокий - спонгиозный губчатый, поверхностный - компактный. Железы расширяются, стенки их становятся складчатыми; в просвете желез появляется секрет, содержащий гликоген и кислые мукополисахариды. Строма с явлениями периваскулярной децидуальной реакции. Спиральные артерии резко извиты, образуют «клубки» (наиболее достоверный признак, определяющий лютеинизирующий эффект). Структура и функциональное состояние эндометрия на 20-22 день 28-дневного менструального цикла представляют оптимальные условия для имплантации бластоцисты.
Поздняя стадия секреции (24-27 день менструального цикла): в указанный период наблюдаются процессы, связанные с регрессом желтого тела и, следовательно, снижением концентрации продуцируемых им гормонов - нарушается трофика эндометрия, формируются дегенеративные его изменения, морфологически эндометрий регрессирует, появляются признаки его ишемии. При этом уменьшается сочность ткани, что приводит к сморщиванию стромы функционального слоя. Складчатость стенок желез усиливается. На 26-27 день менструального цикла в поверхностных слоях компактного слоя наблюдаются лакунарные расширения капилляров и очаговые кровоизлияния в строму; вследствие расплавления волокнистых структур появляются участки разъединения клеток стромы и эпителия желез. Подобное состояние эндометрия именуется «анатомической менструацией» и непосредственно предшествует клинической менструации.
Фаза кровотечения, десквамация (28-2 день менструального цикла). В механизме менструального кровотечения ведущее значение отводится нарушениям кровообращения, обусловленным длительным спазмом артерий (стаз, образование тромбов, ломкость и проницаемость сосудистой стенки, кровоизлияние в строму, лейкоцитарная инфильтрация). Итогом этих преобразований являются некробиоз ткани и ее расплавление. Вследствие расширения сосудов, наступающего после длительного спазма, в ткань эндометрия поступает большое количество крови, что приводит к разрыву сосудов и отторжению - десквамации - некротизированных отделов функционального слоя эндометрия, т. е. к менструальному кровотечению.
Фаза регенерации (3-4 день менструального цикла) короткая, характеризуется регенерацией эндометрия из клеток базального слоя. Эпителизация раневой поверхности происходит из краевых отделов желез базального слоя, а также из неотторгнувшихся глубоких отделов функционального слоя.
Маточные трубы
Функциональное состояние маточных труб варьирует в зависимости от фазы менструального цикла. Так, в лютеиновой фазе цикла активизируется реснитчатый аппарат мерцательного эпителия, возрастает высота его клеток, над апикальной частью которых скапливается секрет. Изменяется также тонус мышечного слоя труб: к моменту овуляции регистрируются урежение и усиление их сокращений, имеющих как маятниковый, так и вращательно-поступательный характер.
Примечательно, что мышечная активность неравнозначна в различных отделах органа: перистальтические волны более характерны для дистальных отделов. Активизация реснитчатого аппарата мерцательного эпителия, лабильность мышечного тонуса маточных труб в лютеиновой фазе, асинхронизм и разнохарактерность сократительной активности в различных отделах органа в совокупности детерминированы для обеспечения оптимальных условий транспорта гамет.
Кроме этого в различные фазы менструального цикла изменяется характер микроциркуляции маточных труб. В периоде овуляции вены, кольцеобразно охватывающие воронку и проникающие вглубь ее бахромок, переполняются кровью, вследствие этого усиливается тонус фимбрий и воронка, приближаясь к яичнику, охватывает его, что параллельно с другими механизмами обеспечивает поступление в трубу овулировавшей яйцеклетки. При прекращении застоя крови в кольцевых венах воронки последняя отодвигается от поверхности яичника.
Влагалище
В течении менструального цикла структура эпителия влагалища претерпевает изменения, соответствующие пролиферативной и регрессивной фазам.
Пролиферативная фаза соответствует фолликулиновой стадии яичников и характеризуется разрастанием, укрупнением и дифференциацией эпителиальных клеток. В период, соответствующий ранней фолликулиновой фазе, разрастание эпителия происходит, главным образом, за счет клеток базального слоя, к середине фазы увеличивается содержание промежуточных клеток. В предовуляторном периоде, когда эпителий влагалища достигает максимальной толщины - 150-300 мкм - наблюдается активация клеток поверхностного слоя: клетки увеличиваются в размерах, ядро их уменьшается, становится пикнотическим. В указанный период возрастает содержание гликогена в клетках базального и, особенно, промежуточного слоев. Отторгаются только единичные клетки.
Регрессивная фаза соответствует лютеиновой стадии. В этой фазе разрастание эпителия прекращается, толщина его уменьшается, часть клеток подвергается обратному развитию. Заканчивается фаза десквамацией клеток большими и компактными группами.
Избранные лекции по акушерству и гинекологии
Под ред. А.Н. Стрижакова, А.И. Давыдова, Л.Д. Белоцерковцевой
Регуляция репродуктивной функции осуществляется единой функциональной нейроэндокринной системой. Функциональная система должна обязательно включать в себя центральное - интегрирующее звено и периферические (эффекторные, исполнительные) органы с различным количеством промежуточных звеньев (рис. 14).
Рис. 14.
В нейроэндокринной системе регуляции репродуктивной функции выделяют пять звеньев, взаимодействующих между собой по принципу прямой и обратной отрицательной и положительной взаимосвязи, определяемой характером сигналов, поступающих с периферии. Ведущая роль в деятельности нейроэндокринных систем отводится отрицательным обратным связям. Физиология и патология менструального цикла в клиническом аспекте наиболее полно отражают состояние репродуктивной системы женщины.
Высшим уровнем регуляции репродуктивной системы являются структуры, составляющие акцептор результата действия. В них воспринимается результат действия всей системы и ее интеграция. К структурам акцептора результата действия относятся высшие отделы нервной системы, миндалевидный комплекс, гиппокамп и другие гипоталамические структуры Они влияют на функцию гипоталамуса и гипофиза, причем влияние может быть стимулирующим и ингибирующим. Различные участки указанных структур стимулируют или тормозят секрецию и выброс гонадолиберинов и гонадотропинов, ускоряют или блокируют овуляцию, ускоряют или задерживают половое развитие, повышают или снижают сексуальность. Физиологические эффекты структур высшего уровня регуляции осуществляются через нервные и гуморальные связи. Ведущее место в этих взаимосвязях отводится мозговым нейротрансмиттерам (катехоламины, серотонин, ацетилхолин, ГАМК, глютаминовая кислота, энкефалины), которые идентефицированы в экстрапирамидных образованиях. Церебральные нейротрансмиттеры регулируют гипоталамо-гипофизарно-яичниковый уровни репродуктивной функции. Так, они определяют циркадные и цирхоральные ритмы, которые являются ведущими в функционировании всей репродуктивной системы. Эндокринный гомеостаз организма поддерживается гипоталамо-гипофизарной системой, состояние которой регулируется нейротрансмиттерами. Основными из них являются биогенные амины и энкефалины.
Биогенные амины, в частности катехоламины (адреналин, норадреналин, дофамин) и серотонин, - высокоэффективные физиологические вещества, как нейромедиаторы и гормоны оказывают многогранное влияние на репродуктивную систему и на все органы и системы организма в физиологических и патологических условиях.
Открытие эндогенных опиоидных пептидов (ЭОП) в середине 1970-х годов позволило значительно расширить представления о нейроэндокринной системе регуляции репродуктивной функции. Выделяют три группы ЭОП: энкефалины, эндорфины и динорфины. ЭОП имеются в различных структурах мозга, вегетативной нервной системы, других тканях и органах (поджелудочной железе, ЖКТ, плаценте, сердце, желчном пузыре и др.), а также в биологических жидкостях организма (плазме, амниотической, фолликулярной, сперме и др.). ЭОП влияют на созревание фолликула, желтого тела, овуляцию, синтез и выброс гормонов гипоталамусом и гипофизом, а это свидетельствует об их многообразной роли в регуляции репродуктивной системы и участии в формировании различных ее патологических состояний. Так, с повышением уровня ЭОП связывается возникновение различных форм аменорей центрального генеза, синдрома поликистозных яичников, болезни Иценко-Кушинга, гирсутизма, ожирения. С помощью ингибиторов опиоидных рецепторов (ОР) налоксона и налтрексона устраняются угнетающие эффекты ЭОП на функцию гипофиза, овуляцию и другие процессы генеративной функции. Установлены и многие другие эффекты ЭОП: терморегуляция (способствуют гипертермии) и антиноцептивное (обезболивающее) действие.
В то же время в клинической практике возможности использования ЭОП с целью регуляции репродуктивной системы до настоящего времени весьма ограниченны.
В структуре высшего звена регуляции репродуктивной функции рассматривается и эпифиз как важнейший нейроэндокринный передатчик, ранее известный как ингибитор развития половой системы. Метилиндолы и пептиды, содержащиеся в эпифизе, по месту их биосинтеза и физиологической роли разделяются на три группы: нейрогипофизарные (аргинин - вазопрессин, аргинин - вазотонин, окситоцин, нейрофизин), воспринимающие информацию от мозга; аденогипофизарные (МСГ, ЛГ, ФСГ, СТГ, ПРЛ), передающие информацию о состоянии регуляторной способности эндокринной системы, собственный гормон эпифиза - мелатонин, ингибирующий уровень ЛГ и ПРЛ. Освобождается мелатонин ритмично с учетом времени суток (больше ночью) и сезонов года (больше зимой). Установлена роль эпифиза в регуляции полового созревания, функционального состояния гипоталамуса, гипофиза и яичников, беременности, лактации, а также в развитии ряда гинекологических заболеваний. Следовательно, эпифиз занимает важное место в нейроэндокринном гомеостазе организма.
Вторым уровнем регуляции репродуктивной функции является гипоталамус, в частности его гипофизотропная зона, состоящая из нейронов вентро- и дорсомедиальных аркуатных ядер, обладающих нейросекреторной активностью либеринов и статинов. Все 10 либеринов и статинов гипоталамуса участвуют в регуляции репродуктивной функции, взаимодействуя между собой. Особая роль при этом принадлежит люлиберину, фолиберину и пролактостатину-пролактинингибирующему фактору (ПИФ). Стимулируется выработка ПРЛ тиролиберином. Главная роль в регуляции выделения ПРЛ принадлежит дофаминергическим структурам. Так, дофамин тормозит освобождение пролактина из лактофоров гипофиза, а его антагонисты (метилдофа, резерпин, аминазин) усиливают его выделение. Путем воздействия на адренергические структуры с помощью парлодела (бромкриптина) удается успешно лечить гиперпролактинемию функционального и органического генеза.
Цирхоральная нейросекреторная функция гипоталамуса, которая модулируется импульсами из экстрагипоталамических структур и коры головного мозга, формируется в пубертатном возрасте и свидетельствует о зрелости нейросекреторных структур гипоталамуса. Регулируя гипофизарно-яичниковую подсистему, она запускает генеративную функцию. Следует также отметить, что определенную роль в выделении гипоталамусом гормонов, как и нейротрансмиттеров, играет уровень эстрадиола в крови.
Третьим уровнем регуляции репродуктивной функции является гипофиз. В нем вырабатываются тропные гормоны периферических эндокринных желез (ФСГ, ЛГ, ПРЛ, ТТГ, АКТГ) и другие. Гонадотропины, взаимодействуя между собой, оказывают влияние на функцию яичников. ФСГ стимулирует рост и созревание фолликулов, секрецию ими эстрогенов. Образование и деятельность желтого тела контролируются ЛГ и ПРЛ. ПРЛ также контролирует рост молочных желез и процесс лактации. В то же время эстрогены ингибируют синтез и выброс ФСГ, а прогестерон - ЛГ и ПРЛ. Таким образом, в зависимости от концентрации и соотношения половых стероидных гормонов угнетается или активируется продукция соответствующих тропных гормонов гипофиза.
Периферические эндокринные органы (яичники, щитовидная железа, надпочечники) представляют четвертый уровень регуляции репродуктивной функции. Главная роль в них принадлежит яичникам.
В них происходят процессы биосинтеза стероидов и развития фолликулов. Фолликулогенез начинается в антенатальном периоде, заканчивается в постменопаузальном. Большинство фолликулов претерпевают атретические изменения и только часть (до 10%) проходит полный цикл развития от примордиального до преовуляторного, затем после овуляции превращается в желтое тело. Доминантный фолликул в первые дни менструального цикла имеет диаметр 2 мм, к моменту овуляции увеличивается до 20-25 мм. Количество фолликулярной жидкости к овуляции увеличивается в 100 раз и более, а клетки гранулезы - до 50 х 106 с 0,5 х 106. В фолликулярной жидкости резко возрастает уровень гормонов, особенно эстрадиола и ФСГ, за счет чего отмечается пик ЛГ и происходит овуляция - разрыв базальной мембраны доминантного фолликула и кровотечение из капилляров тека-клеток. Процесс овуляции совершается с участием простагландинов (F2 и Е2), протеолитических ферментов, окситоцина и релаксина. На процесс овуляции влияют и внешние факторы (питание, стрессовые ситуации, световой и температурный режимы), но главными являются половые гормоны. Повышается уровень люлиберина, затем на фоне увеличенной секреции эстрадиола и последующего пика ЛГ при повышенной чувствительности гипофиза к люлиберину и происходит овуляция. Накануне ее имело место снижение уровня ПРЛ. Затем начинается следующая фаза цикла - лютеиновая, или фаза желтого тела. Овуляция может сопровождаться кратковременными болями внизу живота. Вскоре увеличиваются слизистые выделения из влагалища, происходят падение базальной температуры с повышением ее уже на следующий день, увеличение уровня прогестерона и секреторная трансформация эндометрия, а также другие изменения в различных органах и системах организма. Все это лежит в основе методов диагностики овуляции и ряда патологических состояний - тестов функциональной диагностики.
Развитие фолликулов с созреванием яйцеклетки и в последующем желтого тела происходит одновременно с интенсивным процессом биосинтеза стероидных половых гормонов - эстрогенов, прогестерона и андрогенов. В первую фазу цикла в яичнике эстрадиола секретируется 50- 100 мкг/сут, прогестерона - 2-5 мг/сут, во вторую фазу - соответственно 200-300 мкг/сут и 20- 25 мг/сут, а к моменту овуляции - 400-900 мкг/сут эстрадиола и 10-15 мг/сут прогестерона. В яичниках синтезируются и андрогены (андростендион) до 1,5 мг/сут (или 0,15 мг/сут тестостерона). Столько же его образуется и в надпочечниках. В меньшем количестве тестостерон секретируется в яичниках и в большем - в коре надпочечников из дегидроэпиандростерона и дегидроэпиандростерона сульфата. Тестостерон путем ароматизации с помощью энзимов превращается в дегидротестостерон - наиболее активный андроген, количество которого в женском организме составляет 50-75 мкг/сут. Андрогены синтезируются в тека-клетках, а эстрогены - в гранулезных клетках, в том числе из андрогенов, поступающих из тека-клеток
В гранулезных клетках фолликулов образуются также ингибин. тормозящий выделение ФСГ гипофизом, белковые вещества местного действия - окситоцин и релаксин, а также простагландины. Окситоцин оказывает лютеолитическое действие на желтое тело, а релаксин - токолитическое на миометрий
Максимальная гормональная активность структур всех четырех уровней отмечается в периовуляторный период (до, во время и после овуляции). Неоднозначные, а иногда и противоречивые данные о локализации и содержании рилизинг-гормонов, как и гормонов нижерасположенных структур, свидетельствуют о том, что функциональное состояние всех уровней регуляции следует рассматривать только в тесной взаимосвязи. Так, секреторная функция гипоталамуса по продукции рилизинг-гормонов зависит не только от нейротрансмиттерных механизмов, но и в значительной степени от информации, поступающей с периферии, об уровне циркулирующих в крови гормонов, от скорости их утилизации, что связано с активностью инактивирующих их ферментов. Поэтому наличие в гипоталамических нейроцитах и аденоцитах гипофиза эстрогенных рецепторов (с учетом уровня эстрогенов в крови) не только влияет на продукцию гонадолиберинов, но и модулирует чувствительность к ним у гонадотропинов гипофиза. Подобным же образом уровень стероидных гормонов определяет влияние рилизинг-гормонов на идентичные аденоциты гипофиза. При дефиците периферических гормонов клетки гипофиза становятся высокочувствительными и на аналогичную концентрацию рилизинг-гормона отвечают выбросом большого количества гонадотропинов. Нейротрансмиттерные механизмы (с учетом уровня гормонов в крови) регулируют не только функцию гипоталамуса, но и осуществляют контроль за тройными гормонами гипофиза.
Влияние надпочечников на регуляцию репродуктивной функции осуществляется через гормоны коры и мозгового вещества этих желез. Кора надпочечников имеет морфологическое сходство с корой яичников по происхождению из мезодермальных зачатков, что и определяет сходство их гормонов по химическому строению, а также по цикличности биосинтеза. Известно, что различные нарушения биосинтеза и метаболизма кортикостероидов приводят к патологии гормональной функции яичников и обусловливают многие гинекологические заболевания (синдром Иценко-Кушинга, склерополикистозных яичников и др.). С другой стороны, всевозможные нарушения генеративной функции сопровождаются патофизиологическими изменениями коры надпочечников. Не менее очевидна роль симпатоадреналовой системы в общей НЭС регуляции репродуктивной функции. С помощью катехоламинов можно воздействовать на процессы созревания фолликулов, желтого тела и овуляции.
Влиянием на адренергические структуры с помощью парлодела (бромкриптина) удается корригировать нарушения генеративной функции, наблюдающиеся при гиперпролактинемическом синдроме. Установлена также взаимосвязь структур регуляции функции яичников и надпочечников в гипоталамусе и гипофизе, а также через нейротрансмиттерные механизмы.
Роль щитовидной железы в регуляции репродуктивной функции отчетливо просматривается как в норме, так и особенно при ее функциональных нарушениях по типу гипер- и гипотиреоза. Избыток Т3 и Т4 приводит к увеличению ЛГ, подавлению овуляторного пика гормонов, недостаточности лютеиновой фазы, нарушениям менструального цикла и бесплодию. При дефиците тиреоидных гормонов снижается биосинтез ФСГ и ЛГ, угнетается функция яичников со всеми дальнейшими проявлениями их недостаточности.
Патология гипофизарно-тиреоидной системы особенно отражается на течении беременности, развитии плода. Если выраженные клинические формы заболеваний щитовидной железы вызывают различные нарушения менструального цикла и бесплодие, то при субклинических формах наблюдаются невынашивание и другие осложнения в течении беременности и развитии плода.
Пятым уровнем регуляции репродуктивной системы являются половые органы и молочные железы женщины, а также кожа, кости и жировая ткань. В них реализуют свое действие половые стероидные гормоны, в связи с чем эти органы являются органами-мишенями. Клетки этих тканей и органов имеют рецепторы половых гормонов. Выделяют цитоплазменные (цитозолрецепторы) и ядерные рецепторы. Цитоплазменные рецепторы строго специфичны к эстрогенам, прогестерону и тестостерону, а ядерные могут быть акцепторами (наряду со стероидными гормонами) аминопептидов, инсулина и глюкагона. Для прогестерона по рецепторному связыванию антагонистами считаются глюкокортикоиды.
Следует отметить, что, по мнению П.В Сергеева (1984,1987), первой структурой, взаимодействующей со стероидными гормонами в процессе реализации биологической активности на клеточном уровне, является не цитоплазма, а цитоплазматическая мембрана клеток-мишеней, в которой также содержатся рецепторы стероидных гормонов.
Рецепторы к половым гормонам обнаружены во всех структурах репродуктивной системы и в центральной нервной системе. Содержание стероидных рецепторов и их активность в половых органах, и особенно в эндометрии, изменяются с учетом фазы цикла, т.е. зависят от уровня гормонов в крови.
Рецепторы в клетках яичников (к гонадотропинам), гипофиза (к рилизинг-гормонам) и гипоталамуса (к нейротрансмиттерам) локализуются на клеточной мембране.
Гормон-рецепторные комплексы взаимодействуют с различными структурами клетки-мишени и посредством метаболических реакций проявляют конечные гормональные эффекты. Благодаря им и совершаются многочисленные изменения в половых органах и во всех структурах репродуктивной системы, которые характеризуются по периодам жизни женщины при различных физиологических и патологических состояниях.
Функционирование клеточных рецепторных систем генетически детерминированно, что и определяет как время полового созревания, так и продолжительность периодов жизни женщины и деятельности ее репродуктивной системы.
Характеризуя нейроэндокринную систему регуляции репродуктивной функции женщины, важно учитывать биоритмы (суточные, месячные, годовые, сезонные, многолетние и т.д.). Сущность сводится к тому, что колебания различных физиологических процессов, в том числе и генеративной функции, различаются по указанным периодам. Фазовая временная организация различных функций организма обеспечивается также регуляторными органами и системами. Многие суточные ритмы колебаний выделения гормонов связаны с деятельностью эпифиза, циркадные определяются гипоталамусом и т.д. Нарушение привычного режима жизни при стрессовых ситуациях, резких изменениях климатических условий приводит к состоянию десинхроноза. Хотя эндокринные функции относятся к инертным системам, десинхронозы возможны и в них. Это может привести к различным нарушениям репродуктивной способности и гинекологическим заболеваниям.
В заключение можно отметить высокую надежность и приспособляемость нейроэндокринной регуляции репродуктивной системы к постоянно меняющимся условиям внешней среды за счет многосторонних адаптационных механизмов.